Le développement des membres chez les vertébrés est un domaine de recherche actif en biologie du développement et de l'évolution , une grande partie de ces derniers travaux étant axés sur la transition de la nageoire au membre .
La formation des membres commence dans le champ morphogénétique des membres , lorsque les cellules mésenchymateuses du mésoderme de la plaque latérale prolifèrent au point de provoquer le gonflement de l' ectoderme situé au-dessus, formant ainsi un bourgeon de membre . Le facteur de croissance des fibroblastes (FGF) induit la formation d'un organisateur à l'extrémité du bourgeon de membre, appelé crête ectodermique apicale (AER), qui guide le développement ultérieur et contrôle la mort cellulaire . La mort cellulaire programmée est nécessaire pour éliminer les sangles entre les doigts .
Le champ des membres est une région spécifiée par l'expression de certains gènes Hox , un sous-ensemble de gènes homéotiques et des facteurs de transcription T-box – Tbx5 pour le développement des membres antérieurs ou des ailes, et Tbx4 pour le développement des pattes ou des membres postérieurs. L'établissement du champ des membres antérieurs (mais pas du champ des membres postérieurs) nécessite une signalisation d'acide rétinoïque dans le tronc en développement de l'embryon d'où émergent les bourgeons des membres. De plus, bien que l'excès d'acide rétinoïque puisse modifier la configuration des membres en activant de manière ectopique l'expression de Shh ou Meis1/Meis2, des études génétiques chez la souris qui éliminent la synthèse d'acide rétinoïque ont montré que l'AR n'est pas nécessaire à la configuration des membres.
Le bourgeon du membre reste actif pendant une grande partie du développement du membre car il stimule la création et la rétention de rétroaction positive de deux régions de signalisation : l'AER et sa création ultérieure de la zone d'activité polarisante (ZPA) avec les cellules mésenchymateuses . En plus de l'axe dorso-ventral créé par l'expression ectodermique des signaux compétitifs Wnt7a et BMP respectivement, ces centres de signalisation AER et ZPA sont essentiels à la formation appropriée d'un membre correctement orienté avec sa polarité axiale correspondante dans l'organisme en développement. Étant donné que ces systèmes de signalisation soutiennent réciproquement l'activité de l'autre, le développement du membre est essentiellement autonome après l'établissement de ces régions de signalisation.
Formation des membres
Bourgeon de membre
La formation des membres commence dans le champ morphogénétique des membres . Elle résulte d'une série d'interactions tissulaires réciproques entre le mésenchyme du mésoderme de la plaque latérale et les cellules épithéliales sus-jacentes dérivées de l'ectoderme . Les cellules du mésoderme de la plaque latérale et du myotome migrent vers le champ des membres et prolifèrent au point de provoquer le gonflement de l' ectoderme situé au-dessus, formant ainsi le bourgeon du membre . Les cellules de la plaque latérale produisent les parties cartilagineuses et squelettiques du membre tandis que les cellules du myotome produisent les composants musculaires .
Les cellules mésodermiques de la plaque latérale sécrètent des facteurs de croissance des fibroblastes ( FGF7 et FGF10 ) pour inciter l' ectoderme sus-jacent à former un organisateur à l'extrémité du bourgeon du membre, appelé crête ectodermique apicale (AER), qui guide le développement ultérieur et contrôle la mort cellulaire . L'AER sécrète d'autres facteurs de croissance FGF8 et FGF4 qui maintiennent le signal FGF10 et induisent la prolifération dans le mésoderme. La position de l'expression de FGF10 est régulée par deux voies de signalisation Wnt : Wnt8c dans le membre postérieur et Wnt2b dans le membre antérieur. Le membre antérieur et le membre postérieur sont spécifiés par leur position le long de l' axe antérieur/postérieur et éventuellement par deux facteurs de transcription : Tbx5 et Tbx4 , respectivement.
Condensations précartilagineuses
Les éléments squelettiques des membres sont préfigurés par des agrégats serrés appelés condensations cellulaires des cellules mésenchymateuses précartilagineuses. La condensation mésenchymateuse est médiée par la matrice extracellulaire et les molécules d'adhésion cellulaire . Dans le processus de chondrogenèse , les chondrocytes se différencient des condensations pour former du cartilage , donnant naissance aux primordiums squelettiques . Dans le développement de la plupart des membres des vertébrés (mais pas chez certains amphibiens), le squelette cartilagineux est remplacé par de l'os plus tard au cours du développement.
Périodicités du motif des membres
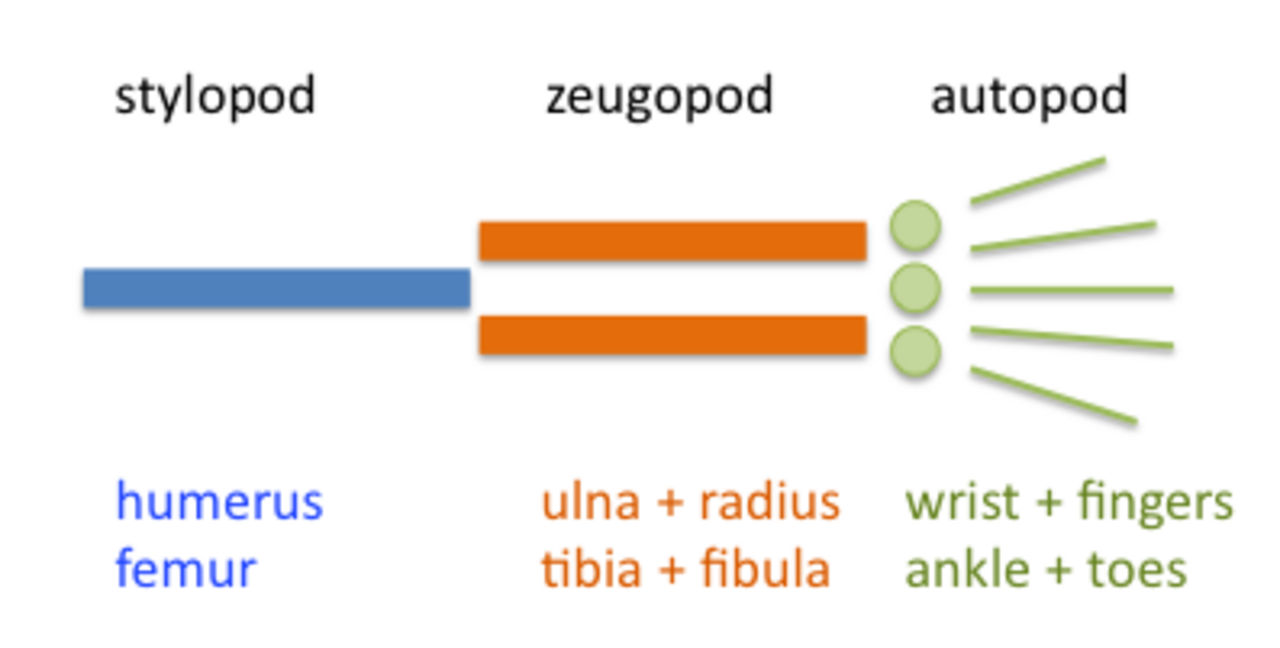
Le membre est organisé en trois régions : stylopode , zeugopode et autopode (dans l'ordre du proximal au distal ). Le zeugopode et l'autopode contiennent un certain nombre de motifs périodiques et quasi-périodiques . Le zeugopode est constitué de deux éléments parallèles le long de l'axe antéro-postérieur et l'autopode contient trois à cinq éléments (dans la plupart des cas) le long du même axe. Les doigts ont également une disposition quasi-périodique le long de l'axe proximodistal, constituée de chaînes en tandem d'éléments squelettiques. La génération du plan de base du membre au cours du développement résulte de la structuration du mésenchyme par une interaction de facteurs qui favorisent la condensation du précartilage et de facteurs qui l'inhibent.
Le développement du plan de base du membre s'accompagne de la génération de différences locales entre les éléments. Par exemple, le radius et l'ulna du membre antérieur, ainsi que le tibia et le péroné du membre postérieur du zeugopode sont distincts les uns des autres, de même que les différents doigts ou orteils de l'autopode. Ces différences peuvent être traitées schématiquement en considérant comment elles se reflètent dans chacun des trois axes principaux du membre.
Il existe un consensus général selon lequel la structuration du squelette des membres implique un ou plusieurs mécanismes de réaction-diffusion de type Turing .
Évolution et développement
L'évolution des membres à partir de nageoires appariées a fait l'objet de nombreuses recherches. De nombreuses études ont été menées pour élucider les gènes et les facteurs de transcription spécifiques responsables du développement des membres (voir le tableau 1 ci-dessous). En particulier, des études ont montré que SHH , DACH1 et les familles de gènes BMP , HOX , T-box , FGF et WNT jouent tous un rôle essentiel dans le développement et la formation réussis des membres. Afin d'étudier les gènes impliqués dans le développement des membres (et donc dans l'évolution), la réduction et la perte des membres chez les serpents constituent une approche complémentaire. Les séquences conservées impliquées dans le développement des membres sont conservées dans les génomes des serpents. Certaines séquences renforçatrices des membres sont également conservées entre différents types d'appendices, tels que les membres et le phallus . Par exemple, la signalisation du développement des membres joue un rôle à la fois dans le développement des membres et du tubercule génital chez la souris. L'étude de la réduction et de la perte des membres révèle les voies génétiques qui contrôlent le développement des membres. Le système de Turing a permis un certain nombre de résultats possibles dans les étapes évolutives des réseaux de structuration.
De nombreux gènes répertoriés dans le tableau 1 jouent un rôle important dans le développement embryonnaire, en particulier lors de la formation du squelette et de la formation des bourgeons des membres. Le gène Shh et les gènes appartenant aux familles BMP, Hox, T-box, FGF et Wnt jouent tous un rôle essentiel dans la signalisation et la différenciation cellulaires pour réguler et favoriser la formation réussie des membres. Divers autres gènes énumérés ci-dessus, dont Dach1, sont des protéines de liaison à l'ADN qui régulent l'expression des gènes. La combinaison complexe de l'expression, de la régulation, de l'activation et de la désactivation des gènes permet à ces gènes de produire des membres pendant le développement embryonnaire. Il est intéressant de noter que bon nombre de ces gènes restent présents même chez les animaux qui n'ont pas de membres, comme les serpents.
Évolution du serpent et perte de membres

Un aspect intéressant de la compréhension du développement des membres est de se demander comment les serpents ont perdu leurs pattes. Les serpents sont un exemple particulièrement pertinent pour étudier la perte des membres, car ils ont subi la perte et la régénération de membres à plusieurs reprises au cours de leur évolution avant de finalement perdre leurs pattes pour de bon. Une grande partie de l'expression des gènes au cours du développement embryonnaire est régulée via la signalisation spatiotemporelle et chimiotactique , comme le montre l'image de droite. Des preuves récentes suggèrent que les gènes hautement conservés responsables du développement des membres (tableau 1) restent présents chez les vertébrés sans membres, indiquant que pendant le développement embryonnaire, la production de membres, ou son absence, peut être mieux expliquée par la régulation des gènes.
Rôle possible de l'amplificateur Shh dans la perte des membres du serpent
Une théorie suggère que la dégradation des séquences amplificatrices pourrait avoir contribué à la perte progressive des membres au cours de l'évolution des serpents . En particulier, de nombreuses études se sont concentrées sur la séquence régulatrice ZPA (ZRS) - l'amplificateur du gène Sonic Hedgehog ( Shh ). Cet amplificateur à longue portée est nécessaire à la formation correcte des membres chez plusieurs espèces de vertébrés , les mutations de cette séquence étant connues pour provoquer des déformations des membres. En tant que telle, cette séquence est hautement conservée dans de nombreuses espèces de vertébrés.
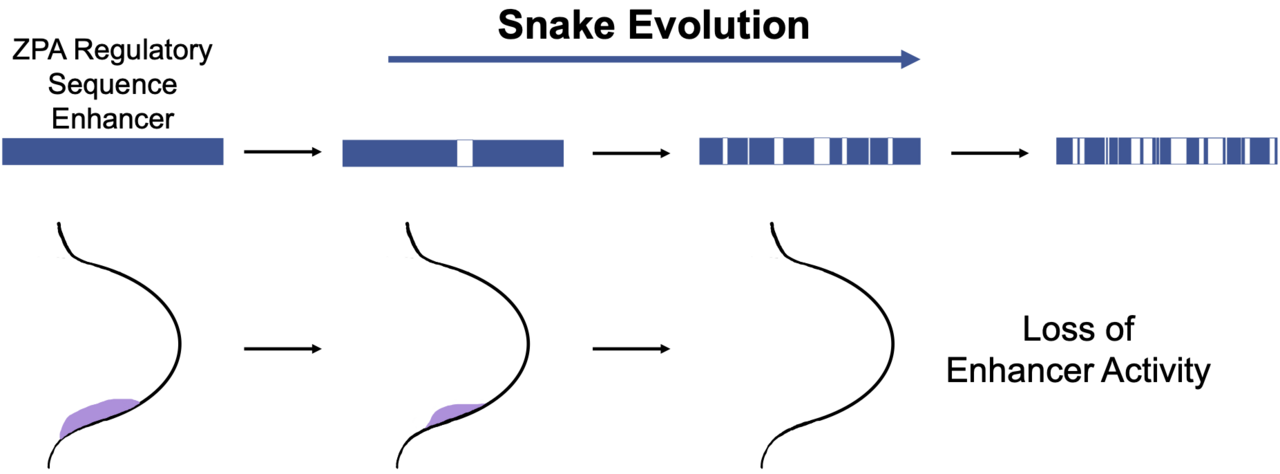
Les comparaisons entre le ZRS de base de plusieurs espèces de serpents et les séquences de souris et de lézards montrent la présence d'altérations spécifiques aux serpents. Le ZRS de base s'est avéré principalement conservé chez les serpents basilaires comme le boa constrictor et le python , qui contiennent encore des os de la ceinture pelvienne . En revanche, les serpents avancés tels que la vipère et le cobra , chez lesquels aucune structure squelettique des membres ne subsiste, ont un taux de changements de nucléotides beaucoup plus élevé par rapport au ZRS de la souris et du lézard. On pense que ces changements cumulatifs dans le ZRS du serpent indiquent une perte progressive de fonction de cet activateur tout au long de l'évolution du serpent.

Des recherches plus poussées sur ces changements ont montré un taux accru de substitution dans les sites de liaison des facteurs de transcription tels que ETS1 , dont la liaison à ZRS s'est avérée activer la transcription de Shh. Cette dégradation dans ZRS suggère que cet activateur pourrait être important pour explorer davantage les mécanismes moléculaires qui ont propulsé l'évolution morphologique des serpents.
Conclusions et limites actuelles
Les serpents ne sont pas un organisme modèle courant, c'est-à-dire qu'ils ne sont pas facilement génétiquement traitables. De plus, les données de séquence de leur génome sont incomplètes et souffrent d'une mauvaise annotation et d'une mauvaise qualité. Ces facteurs rendent difficile la compréhension du mécanisme de la perte des membres des serpents en utilisant une approche génétique, en ciblant et en observant la présence et l'activité de ces gènes et de leurs activateurs régulateurs. De nombreux gènes nécessaires à la formation des membres sont toujours conservés chez les serpents, de sorte que la perte des membres ne peut probablement pas être expliquée par la perte de gènes.
Motifs axiaux et problèmes connexes
Le membre en développement doit s'aligner par rapport à trois axes de symétrie. Il s'agit des axes cranio-caudal (de la tête à la queue), dorso-ventral (de l'arrière vers l'avant) et proximodistal (de près vers l'arrière).
De nombreuses recherches sur le développement du modèle squelettique des membres ont été influencées par le concept d'information positionnelle proposé par Lewis Wolpert en 1971. Conformément à cette idée, des efforts ont été faits pour identifier les molécules de signalisation diffusives ( morphogènes ) qui traversent les axes orthogonaux des membres en développement et déterminent les emplacements et les identités des éléments squelettiques de manière dépendante de la concentration.
Schéma proximodistal
Les gènes Hox contribuent à la spécification du stylopode , du zeugopode et de l'autopode . Les mutations des gènes Hox entraînent des pertes ou des anomalies proximodistales . Trois modèles différents ont été avancés pour expliquer la configuration de ces régions.
Modèle de zone de progression
La crête ectodermique apicale (AER) crée et maintient une zone de prolifération cellulaire appelée zone de progression . On pense que les cellules y obtiennent les informations de position dont elles ont besoin pour se déplacer vers leur position de destination. Il a été proposé que leur valeur de position était déterminée par la durée pendant laquelle les cellules étaient dans la zone de progression, mais cela n'a pas encore été prouvé (en 2001). Il a été proposé que les structures proximales soient formées par les premières cellules à quitter la zone et les structures distales, par les cellules qui sont parties plus tard.
Le modèle de zone de progrès a été proposé il y a 30 ans, mais des preuves récentes sont en contradiction avec ce modèle.
Preuves expérimentales :
- Le retrait de l’AER à une période ultérieure du développement entraîne moins de perturbations des structures distales que si l’AER était retiré au début du développement.
- La greffe d'une extrémité de bourgeon de membre précoce sur une aile tardive entraîne une duplication des structures, tandis que la greffe d'une extrémité de bourgeon d'aile tardif sur un membre précoce entraîne une suppression des structures.
Modèle d'allocation précoce et d'expansion des progéniteurs (ou modèle de pré-spécification)
Les cellules sont spécifiées pour chaque segment du bourgeon de membre précoce et cette population de cellules s'étend à mesure que le bourgeon de membre grandit. Ce modèle est cohérent avec les observations suivantes. La division cellulaire est observée dans tout le bourgeon de membre. La mort cellulaire se produit dans une zone de 200 μm sous-jacente à l'AER lorsqu'il est retiré ; la mort cellulaire empêche une certaine formation de motifs. Les billes libérant du FGF sont capables de sauver le développement du membre lorsque l'AER est retiré en empêchant cette mort cellulaire.
Preuves expérimentales :
- Les cellules marquées dans différentes positions d'un bourgeon de membre précoce étaient limitées à des segments uniques du membre.
- Les membres dépourvus d'expression des FGF4 et FGF8 requis ont montré toutes les structures du membre et pas seulement les parties proximales.
Plus récemment, cependant, les chercheurs principalement responsables des modèles de zone de progrès et de pré-spécification ont reconnu qu'aucun de ces modèles ne rendait compte de manière adéquate des données expérimentales disponibles.
Modèle de réaction-diffusion de type Turing
Ce modèle, un modèle de réaction-diffusion proposé pour la première fois en 1979, est basé sur les propriétés d'auto-organisation des milieux excitables décrites par Alan Turing en 1952. Le milieu excitable est le mésenchyme du bourgeon de membre, dans lequel les cellules interagissent par des morphogènes autorégulateurs positifs tels que le facteur de croissance transformant bêta (TGF-β) et les voies de signalisation inhibitrices impliquant le facteur de croissance des fibroblastes (FGF) et Notch . Les axes proximodistaux et cranio-caudaux ne sont pas considérés comme étant spécifiés indépendamment, mais émergent plutôt par des transitions dans le nombre d'éléments parallèles lorsque la zone apicale indifférenciée du bourgeon de membre en croissance subit un remodelage. Ce modèle ne spécifie qu'un modèle « dépouillé ». D'autres facteurs comme le hérisson sonique (Shh) et les protéines Hox, principales molécules d'information dans les autres modèles, sont proposés à la place pour jouer un rôle de réglage fin.
Preuves expérimentales :
- Les cellules mésenchymateuses des membres, lorsqu'elles sont dissociées et cultivées ou réintroduites dans des « coques » ectodermiques, peuvent récapituler les aspects essentiels de la formation des motifs , de la morphogenèse et de la différenciation .
- Les particularités du modèle squelettique des membres chez le mutant Doublefoot de la souris sont les résultats prédits d'un mécanisme de type Turing.
- La réduction progressive des gènes Hox distaux dans un contexte Gli3 -nul entraîne une polydactylie progressivement plus sévère, présentant des doigts plus fins et plus denses, suggérant (avec l'aide de la modélisation informatique) que la dose de gènes Hox distaux module la période ou la longueur d'onde des doigts spécifiée par un mécanisme de type Turing.
Schéma cranio-caudal
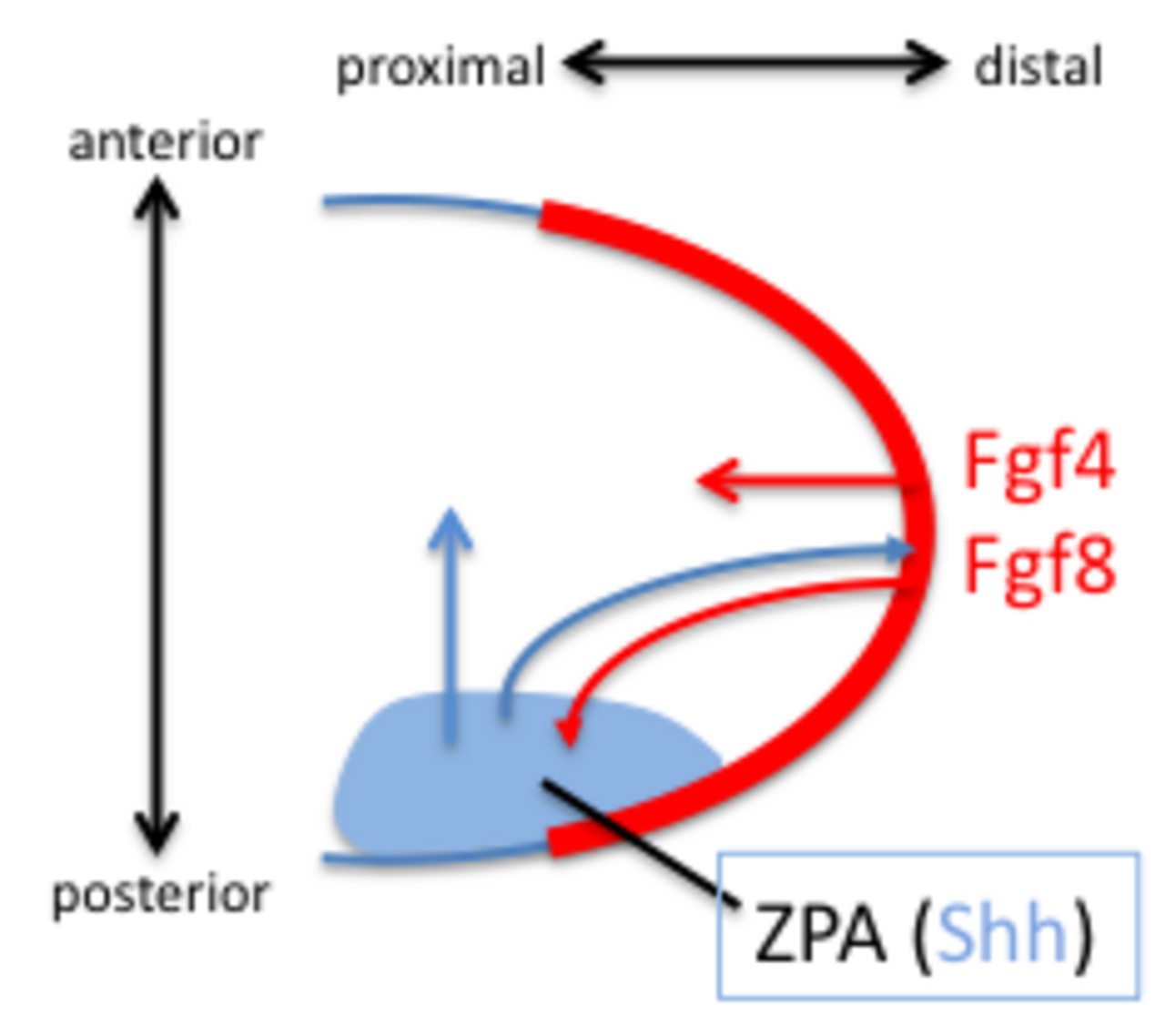
En 1957, la découverte de la zone d'activité polarisante (ZPA) dans le bourgeon du membre a fourni un modèle pour comprendre l'activité de structuration par l'action d'un gradient morphogénique de sonic hedgehog (Shh). Shh est reconnu comme un activateur spécifique du membre. Shh est à la fois suffisant et nécessaire pour créer la ZPA et spécifier le motif cranio-caudal dans le membre distal (Shh n'est pas nécessaire pour la polarité du stylopode). Shh est activé dans la partie postérieure par l'expression précoce des gènes Hoxd, l'expression de Hoxb8 et l'expression dHAND. Shh est maintenu dans la partie postérieure par une boucle de rétroaction entre la ZPA et l'AER. Shh induit l'AER à produire FGF4 et FGF8 qui maintiennent l'expression de Shh.
Les chiffres 3, 4 et 5 sont spécifiés par un gradient temporel de Shh. Le chiffre 2 est spécifié par une forme diffusible à longue portée de Shh et le chiffre 1 ne nécessite pas Shh. Shh clive le complexe répresseur transcriptionnel Ci/Gli3 pour convertir le facteur de transcription Gli3 en un activateur qui active la transcription des gènes HoxD le long du craniocaudal. La perte du répresseur Gli3 conduit à la formation de chiffres génériques (non individualisés) en quantités supplémentaires.
Schéma dorsoventral
La structuration dorsoventrale est médiée par les signaux Wnt7a dans l'ectoderme sus-jacent et non dans le mésoderme. Wnt7a est à la fois nécessaire et suffisant pour dorsaliser le membre. Wnt7a influence également le craniocaudal et la perte de Wnt7a fait que le côté dorsal des membres devient le côté ventral et provoque l'absence de doigts postérieurs. Le remplacement des signaux Wnt7a corrige ce défaut. Wnt7a est également nécessaire pour maintenir l'expression de Shh.
Wnt7a provoque également l'expression de Lmx1b, un gène LIM Homeobox (et donc un facteur de transcription ). Lmx1b est impliqué dans la dorsalisation du membre, ce qui a été démontré par l'inactivation du gène Lmx1b chez la souris. Les souris dépourvues de Lmx1b ont produit de la peau ventrale des deux côtés de leurs pattes. D'autres facteurs semblent contrôler le motif DV ; Engrailed-1 réprime l'effet dorsalisant de Wnt7a sur le côté ventral des membres.