Le système de récompense (le circuit mésocorticolimbique) est un groupe de structures neuronales responsables de la saillance de l'incitation (c'est-à-dire « vouloir » ; désir ou envie d'une récompense et motivation ), de l'apprentissage associatif (principalement le renforcement positif et le conditionnement classique ) et des émotions à valence positive , en particulier celles impliquant le plaisir comme composante principale (par exemple, la joie , l'euphorie et l'extase ). La récompense est la propriété attractive et motivationnelle d'un stimulus qui induit un comportement appétitif, également connu sous le nom de comportement d'approche et de comportement de consommation. Un stimulus gratifiant a été décrit comme « tout stimulus, objet, événement, activité ou situation qui a le potentiel de nous faire l'approcher et le consommer est par définition une récompense ». Dans le conditionnement opérant , les stimuli gratifiants fonctionnent comme des renforçateurs positifs ; cependant, l'affirmation inverse est également vraie : les renforçateurs positifs sont gratifiants. Le système de récompense motive les animaux à s'approcher des stimuli ou à adopter un comportement qui augmente leur condition physique (sexe, aliments riches en énergie, etc.). La survie de la plupart des espèces animales dépend de la maximisation du contact avec des stimuli bénéfiques et de la minimisation du contact avec des stimuli nocifs. La cognition de récompense sert à augmenter la probabilité de survie et de reproduction en provoquant un apprentissage associatif, en suscitant un comportement d'approche et de consommation et en déclenchant des émotions à valence positive . Ainsi, la récompense est un mécanisme qui a évolué pour aider à augmenter la condition physique adaptative des animaux. Dans la toxicomanie , certaines substances suractivent le circuit de récompense, ce qui conduit à un comportement compulsif de recherche de substances résultant de la plasticité synaptique du circuit.
Les récompenses primaires sont une classe de stimuli gratifiants qui facilitent la survie de soi-même et de sa progéniture , et elles comprennent les récompenses homéostatiques (par exemple, la nourriture appétissante ) et reproductives (par exemple, le contact sexuel et l'investissement parental ). Les récompenses intrinsèques sont des récompenses inconditionnelles qui sont attrayantes et motivent le comportement parce qu'elles sont intrinsèquement agréables. Les récompenses extrinsèques (par exemple, l'argent ou voir son équipe sportive préférée gagner un match) sont des récompenses conditionnées qui sont attrayantes et motivent le comportement mais ne sont pas intrinsèquement agréables. Les récompenses extrinsèques tirent leur valeur motivationnelle d'une association apprise (c'est-à-dire un conditionnement) avec des récompenses intrinsèques. Les récompenses extrinsèques peuvent également susciter du plaisir (par exemple, l'euphorie de gagner beaucoup d'argent à la loterie) après avoir été conditionnées de manière classique avec des récompenses intrinsèques.
Définition
En neurosciences, le système de récompense est un ensemble de structures cérébrales et de voies neuronales responsables de la cognition liée à la récompense, notamment l'apprentissage associatif (principalement le conditionnement classique et le renforcement opérant ), la saillance incitative (c'est-à-dire la motivation et le « vouloir », le désir ou l'envie d'une récompense) et les émotions à valence positive , en particulier les émotions qui impliquent le plaisir (c'est-à-dire le « goût » hédonique).
Les termes couramment utilisés pour décrire le comportement lié à la composante « vouloir » ou désir de la récompense comprennent le comportement appétitif, le comportement d'approche, le comportement préparatoire, le comportement instrumental, le comportement anticipatoire et la recherche. Les termes couramment utilisés pour décrire le comportement lié à la composante « aimer » ou plaisir de la récompense comprennent le comportement de consommation et le comportement de prise.
Les trois fonctions principales des récompenses sont leur capacité à :
- produire un apprentissage associatif (c'est-à-dire, le conditionnement classique et le renforcement opérant ) ;
- affecter la prise de décision et induire un comportement d'approche (via l'attribution d' une importance motivationnelle à des stimuli gratifiants) ;
- susciter des émotions à valence positive , en particulier le plaisir.
Neuroanatomie
Aperçu
Les structures cérébrales qui composent le système de récompense sont situées principalement dans la boucle cortico-ganglions de la base-thalamo-corticale ; la partie des ganglions de la base de la boucle pilote l'activité au sein du système de récompense. La plupart des voies qui relient les structures au sein du système de récompense sont les interneurones glutamatergiques , les neurones épineux moyens GABAergiques (MSN) et les neurones de projection dopaminergiques , bien que d'autres types de neurones de projection y contribuent (par exemple, les neurones de projection orexinergiques ). Le système de récompense comprend l' aire tegmentale ventrale , le striatum ventral (c'est-à-dire le noyau accumbens et le tubercule olfactif ), le striatum dorsal (c'est-à-dire le noyau caudé et le putamen ), la substance noire (c'est-à-dire la pars compacta et la pars reticulata ), le cortex préfrontal , le cortex cingulaire antérieur , le cortex insulaire , l'hippocampe , l'hypothalamus (en particulier le noyau orexinergique dans l' hypothalamus latéral ), le thalamus (noyaux multiples), le noyau sous-thalamique , le globus pallidus ( externe et interne ), le pallidum ventral , le noyau parabrachial , l'amygdale et le reste de l' amygdale étendue . Le noyau du raphé dorsal et le cervelet semblent également moduler certaines formes de cognition liée à la récompense (c'est-à-dire l'apprentissage associatif , la saillance motivationnelle et les émotions positives ) et les comportements. Le noyau tegmental latérodorsal (LDT) , le noyau pédonculopontin (PPTg) et l'habenula latérale (LHb) (directement et indirectement via le noyau tegmental rostromédien (RMTg) ) sont également capables d'induire une saillance aversive et une saillance incitative par leurs projections vers l'aire tegmentale ventrale (VTA). Le LDT et le PPTg envoient tous deux des projections glutaminergiques à l'aire tegmentale ventrale qui se synapsent sur les neurones dopaminergiques, qui peuvent tous deux produire une saillance incitative. Le LHb envoie des projections glutaminergiques, dont la majorité se synapsent sur les neurones GABAergiques RMTg qui, à leur tour, entraînent l'inhibition des neurones dopaminergiques de l'aire tegmentale ventrale, bien que certaines projections de LHb se terminent sur les interneurones de l'aire tegmentale ventrale. Ces projections de LHb sont activées à la fois par des stimuli aversifs et par l'absence d'une récompense attendue, et l'excitation du LHb peut induire une aversion.
La plupart des voies de la dopamine (c'est-à-dire les neurones qui utilisent le neurotransmetteur dopamine pour communiquer avec d'autres neurones) qui se projettent hors de l'aire tegmentale ventrale font partie du système de récompense ; dans ces voies, la dopamine agit sur les récepteurs de type D1 ou D2 pour stimuler (de type D1) ou inhiber (de type D2) la production d' AMPc . Les neurones épineux moyens GABAergiques du striatum sont également des composants du système de récompense. Les noyaux de projection glutamatergiques dans le noyau sous-thalamique, le cortex préfrontal, l'hippocampe, le thalamus et l'amygdale se connectent à d'autres parties du système de récompense via les voies du glutamate. Le faisceau médial du prosencéphale , qui est un ensemble de nombreuses voies neuronales qui assurent la récompense de stimulation cérébrale (c'est-à-dire la récompense dérivée de la stimulation électrochimique directe de l' hypothalamus latéral ), est également un composant du système de récompense.
Deux théories existent concernant l'activité du noyau accumbens et la génération d'appréciation et de désir. L'hypothèse d'inhibition (ou d'hyperpolarisation) propose que le noyau accumbens exerce des effets inhibiteurs toniques sur les structures en aval telles que le pallidum ventral, l'hypothalamus ou l'aire tegmentale ventrale, et qu'en inhibant les MSN dans le noyau accumbens (NAcc), ces structures sont excitées, « libérant » un comportement lié à la récompense. Alors que les agonistes des récepteurs GABA sont capables de provoquer à la fois des réactions d'appréciation et de désir dans le noyau accumbens, les apports glutaminergiques provenant de l' amygdale basolatérale , de l'hippocampe ventral et du cortex préfrontal médian peuvent favoriser la saillance incitative. De plus, alors que la plupart des études montrent que les neurones NAcc réduisent les décharges en réponse à la récompense, un certain nombre d'études trouvent la réponse opposée. Cela a conduit à la proposition de l'hypothèse de désinhibition (ou de dépolarisation), qui suggère que l'excitation des neurones NAcc, ou au moins certains sous-ensembles, déterminent le comportement lié à la récompense.
Après près de 50 ans de recherche sur la récompense par stimulation cérébrale, les experts ont certifié que des dizaines de sites du cerveau maintiennent l'auto-stimulation intracrânienne . Ces régions comprennent l'hypothalamus latéral et les faisceaux médiaux du prosencéphale, qui sont particulièrement efficaces. La stimulation à ces endroits active les fibres qui forment les voies ascendantes ; les voies ascendantes comprennent la voie dopaminergique mésolimbique , qui se projette de l'aire tegmentale ventrale jusqu'au noyau accumbens . Il existe plusieurs explications quant à la raison pour laquelle la voie dopaminergique mésolimbique est centrale dans les circuits de médiation de la récompense. Tout d'abord, il y a une augmentation marquée de la libération de dopamine par la voie mésolimbique lorsque les animaux s'engagent dans une auto-stimulation intracrânienne. Deuxièmement, les expériences indiquent systématiquement que la récompense par stimulation cérébrale stimule le renforcement des voies qui sont normalement activées par des récompenses naturelles , et la récompense médicamenteuse ou l'auto-stimulation intracrânienne peuvent exercer une activation plus puissante des mécanismes centraux de récompense parce qu'elles activent le centre de récompense directement plutôt que par l'intermédiaire des nerfs périphériques . Troisièmement, lorsque des animaux reçoivent des drogues addictives ou adoptent des comportements naturellement gratifiants, comme l’alimentation ou l’activité sexuelle, il se produit une libération marquée de dopamine dans le noyau accumbens. Cependant, la dopamine n’est pas le seul composé de récompense dans le cerveau.
Voie clé
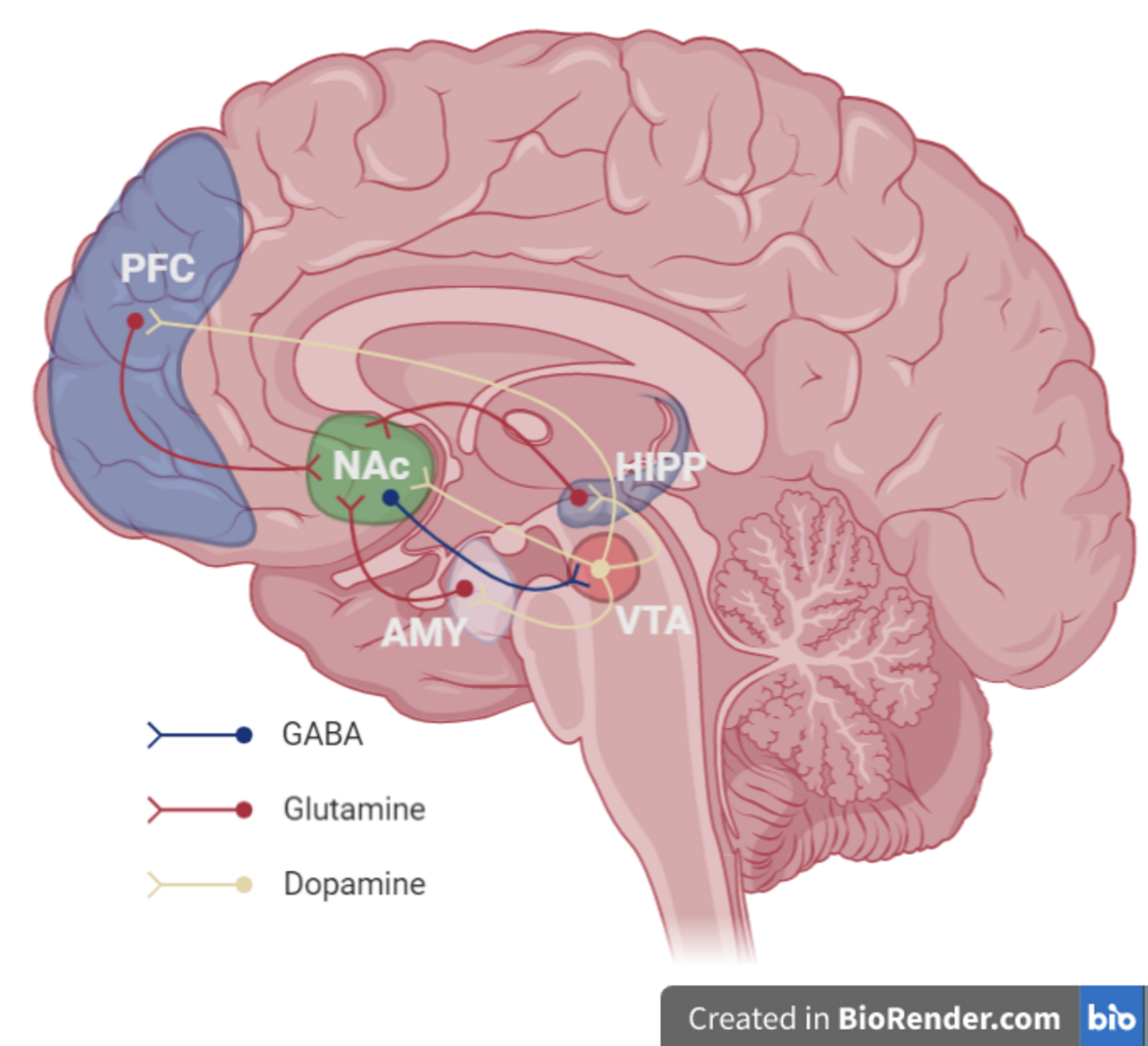
Aire tegmentale ventrale
- L' aire tegmentale ventrale (ATV) joue un rôle important dans la réponse aux stimuli et aux signaux qui indiquent la présence d'une récompense. Les stimuli de récompense (et toutes les drogues addictives) agissent sur le circuit en déclenchant la libération par l'ATV de signaux de dopamine vers le noyau accumbens , directement ou indirectement. L'ATV a deux voies importantes : la voie mésolimbique qui se projette vers les régions limbiques (striatales) et qui sous-tend les comportements et processus motivationnels, et la voie mésocorticale qui se projette vers le cortex préfrontal, qui sous-tend les fonctions cognitives, telles que l'apprentissage de signaux externes, etc.
- Les neurones dopaminergiques de cette région convertissent l'acide aminé tyrosine en DOPA à l'aide de l'enzyme tyrosine hydroxylase , qui est ensuite convertie en dopamine à l'aide de l'enzyme DOPA décarboxylase .
Striatum (Noyau Accumbens)
- Le striatum est largement impliqué dans l'acquisition et la stimulation de comportements appris en réponse à un signal de récompense. Le VTA se projette vers le striatum et active les neurones épineux moyens GABAergiques via les récepteurs D1 et D2 dans le striatum ventral (noyau accumbens) et dorsal.
- Le striatum ventral (le noyau accumbens) est largement impliqué dans l'acquisition du comportement lorsqu'il est alimenté par l'aire tectonique ventrale, et dans la stimulation du comportement lorsqu'il est alimenté par le PFC. La coque NAc se projette sur le pallidum et l'aire tectonique ventrale, régulant les fonctions limbiques et autonomes. Cela module les propriétés de renforcement des stimuli et les aspects à court terme de la récompense. Le noyau NAc se projette sur la substance noire et est impliqué dans le développement des comportements de recherche de récompense et son expression. Il est impliqué dans l'apprentissage spatial, la réponse conditionnelle et le choix impulsif ; les éléments à long terme de la récompense.
- Le striatum dorsal est impliqué dans l'apprentissage, le striatum médial dorsal dans l'apprentissage orienté vers un objectif et le striatum latéral dorsal dans l'apprentissage stimulus-réponse fondamental pour la réponse pavlovienne. Lors d'une activation répétée par un stimulus, le noyau accumbens peut activer le striatum dorsal via une boucle intrastriatale. La transition des signaux du NAc au DS permet aux signaux associés à la récompense d'activer le DS sans que la récompense elle-même ne soit présente. Cela peut activer les envies et les comportements de recherche de récompense (et est responsable du déclenchement de la rechute pendant l'abstinence dans la dépendance).
Cortex préfrontal
- Les neurones dopaminergiques du VTA se projettent vers le PFC, activant les neurones glutaminergiques qui se projettent vers plusieurs autres régions, y compris le striatum dorsal et le NAc, permettant finalement au PFC de servir de médiateur à la saillance et aux comportements conditionnels en réponse aux stimuli.
- Notamment, l'abstinence de drogues addictives active le PFC, une projection glutamatergique vers le NAc, qui conduit à de fortes envies et module la réinstallation des comportements addictifs résultant de l'abstinence. Le PFC interagit également avec le VTA par la voie mésocorticale et aide à associer les signaux environnementaux à la récompense.
Hippocampe
- L'hippocampe a de multiples fonctions, notamment dans la création et le stockage des souvenirs. Dans le circuit de la récompense, il sert aux souvenirs contextuels et aux signaux associés. Il soutient en fin de compte le rétablissement des comportements de recherche de récompense via des signaux et des déclencheurs contextuels.
Amygdale
- L'AMY reçoit des informations de l'ATV et les transmet au NAc. L'amygdale joue un rôle important dans la création de puissants souvenirs émotionnels et soutient probablement la création de souvenirs forts associés à des signaux. Elle joue également un rôle important dans la médiation des effets anxieux du sevrage et de l'augmentation de la consommation de drogues en cas de dépendance.
Centres de plaisir
Le plaisir est une composante de la récompense, mais toutes les récompenses ne sont pas agréables (par exemple, l'argent ne suscite pas de plaisir à moins que cette réponse ne soit conditionnée). Les stimuli qui sont naturellement agréables, et donc attrayants, sont appelés récompenses intrinsèques , tandis que les stimuli qui sont attrayants et motivent un comportement d'approche, mais qui ne sont pas intrinsèquement agréables, sont appelés récompenses extrinsèques . Les récompenses extrinsèques (par exemple, l'argent) sont gratifiantes en raison d'une association apprise avec une récompense intrinsèque. En d'autres termes, les récompenses extrinsèques fonctionnent comme des aimants motivationnels qui suscitent des réactions de « vouloir », mais pas d'« aimer » une fois qu'elles ont été acquises.
Le système de récompense contient des centres de plaisir ou des points chauds hédoniques , c'est-à-dire des structures cérébrales qui médiatisent les réactions de plaisir ou de « goût » à partir de récompenses intrinsèques. En octobre 2017, des points chauds hédoniques ont été identifiés dans des sous-compartiments de la coque du noyau accumbens , du pallidum ventral , du noyau parabrachial , du cortex orbitofrontal (OFC) et du cortex insulaire . Le point chaud de la coque du noyau accumbens est situé dans le quadrant rostro-dorsal de la coque médiale, tandis que le point froid hédonique est situé dans une région plus postérieure. Le pallidum ventral postérieur contient également un point chaud hédonique, tandis que le pallidum ventral antérieur contient un point froid hédonique. Chez les rats, les microinjections d' opioïdes , d'endocannabinoïdes et d'orexine sont capables d'améliorer les réactions d'appréciation dans ces points chauds. Il a été démontré que les points chauds hédoniques situés dans l'OFC antérieur et l'insula postérieure répondent à l'orexine et aux opioïdes chez les rats, tout comme le point froid hédonique superposé dans l'insula antérieure et l'OFC postérieur. D'autre part, il a été démontré que le point chaud du noyau parabrachial ne répond qu'aux agonistes des récepteurs des benzodiazépines.
Les points chauds hédoniques sont fonctionnellement liés, dans la mesure où l'activation d'un point chaud entraîne le recrutement des autres, comme l'indique l' expression induite de c-Fos , un gène précoce immédiat . De plus, l'inhibition d'un point chaud entraîne l'atténuation des effets de l'activation d'un autre point chaud. Par conséquent, l'activation simultanée de chaque point chaud hédonique au sein du système de récompense est considérée comme nécessaire pour générer la sensation d'une euphorie intense .
Vouloir et aimer
La saillance incitative est l'attribut « vouloir » ou « désir », qui comprend une composante motivationnelle, qui est attribué à un stimulus gratifiant par la coque du noyau accumbens (coque NAcc). Le degré de neurotransmission de la dopamine dans la coque NAcc à partir de la voie mésolimbique est fortement corrélé à l'ampleur de la saillance incitative pour les stimuli gratifiants.
L'activation de la région dorsorostrale du noyau accumbens est corrélée à une augmentation du désir sans augmentation simultanée du goût. Cependant, la neurotransmission dopaminergique dans la coque du noyau accumbens est responsable non seulement de la saillance motivationnelle appétitive (c'est-à-dire de la saillance incitative) envers les stimuli gratifiants, mais aussi de la saillance motivationnelle aversive, qui éloigne le comportement des stimuli indésirables. Dans le striatum dorsal, l'activation des MSN exprimant D1 produit une saillance incitative appétitive, tandis que l'activation des MSN exprimant D2 produit de l'aversion. Dans le NAcc, une telle dichotomie n'est pas aussi nette, et l'activation des MSN D1 et D2 est suffisante pour améliorer la motivation, probablement via la désinhibition de l'ATV par l'inhibition du pallidum ventral.
La théorie de la sensibilisation et de l’incitation de Robinson et Berridge, fondée en 1993, suggérait que la récompense contient des composantes psychologiques distinctes : le désir (incitation) et l’appréciation (plaisir). Pour expliquer l’augmentation du contact avec un certain stimulus comme le chocolat, deux facteurs indépendants entrent en jeu : notre désir d’avoir du chocolat (envie) et l’effet de plaisir du chocolat (appréciation). Selon Robinson et Berridge, l’appréciation et l’appréciation sont deux aspects du même processus, de sorte que les récompenses sont généralement désirées et appréciées au même degré. Cependant, l’appréciation et l’appréciation changent également de manière indépendante dans certaines circonstances. Par exemple, les rats qui ne mangent pas après avoir reçu de la dopamine (perte d’appétit) agissent comme s’ils aimaient toujours la nourriture. Dans un autre exemple, des électrodes d’auto-stimulation activées dans l’hypothalamus latéral des rats augmentent l’appétit, mais provoquent également davantage de réactions indésirables aux goûts tels que le sucre et le sel ; apparemment, la stimulation augmente l’appréciation mais pas l’appréciation. De tels résultats démontrent que le système de récompense des rats comprend des processus indépendants d’appréciation et d’appréciation. On pense que la composante désir est contrôlée par les voies dopaminergiques , tandis que la composante goût est contrôlée par les systèmes endocannabinoïdes opiacés-GABA.
Système anti-récompense
Koobs et Le Moal ont suggéré qu'il existe un circuit distinct responsable de l'atténuation du comportement de recherche de récompense, qu'ils ont appelé le circuit anti-récompense. Ce composant agit comme un frein sur le circuit de récompense, empêchant ainsi la recherche excessive de nourriture, de sexe, etc. Ce circuit implique plusieurs parties de l'amygdale (le noyau du lit de la strie terminale, le noyau central), le noyau accumbens et des molécules de signalisation, notamment la noradrénaline, le facteur de libération de la corticotropine et la dynorphine. Ce circuit est également supposé servir de médiateur aux composantes désagréables du stress, et est donc censé être impliqué dans la dépendance et le sevrage. Alors que le circuit de récompense sert de médiateur au renforcement positif initial impliqué dans le développement de la dépendance, c'est le circuit anti-récompense qui domine plus tard via le renforcement négatif qui motive la recherche des stimuli gratifiants.
Apprentissage
Les stimuli récompensants peuvent favoriser l'apprentissage sous la forme de conditionnement classique (conditionnement pavlovien) et de conditionnement opérant (conditionnement instrumental) . Dans le conditionnement classique, une récompense peut agir comme un stimulus inconditionné qui, lorsqu'il est associé au stimulus conditionné, amène le stimulus conditionné à susciter à la fois des réponses musculo-squelettiques (sous la forme de comportements simples d'approche et d'évitement) et végétatives. Dans le conditionnement opérant, une récompense peut agir comme un renforçateur dans la mesure où elle augmente ou soutient les actions qui mènent à elle-même. Les comportements appris peuvent ou non être sensibles à la valeur des résultats auxquels ils conduisent ; les comportements qui sont sensibles à la contingence d'un résultat sur la performance d'une action ainsi qu'à la valeur du résultat sont orientés vers un objectif , tandis que les actions suscitées qui sont insensibles à la contingence ou à la valeur sont appelées habitudes . Cette distinction est censée refléter deux formes d'apprentissage, sans modèle et basé sur un modèle. L'apprentissage sans modèle implique la simple mise en cache et la mise à jour des valeurs. En revanche, l'apprentissage basé sur un modèle implique le stockage et la construction d'un modèle interne d'événements qui permet l'inférence et la prédiction flexible. Bien que le conditionnement pavlovien soit généralement supposé être indépendant du modèle, la saillance incitative attribuée à un stimulus conditionné est flexible en ce qui concerne les changements dans les états motivationnels internes.
Des systèmes neuronaux distincts sont responsables de l'apprentissage des associations entre les stimuli et les résultats, les actions et les résultats, et les stimuli et les réponses. Bien que le conditionnement classique ne se limite pas au système de récompense, l'amélioration de la performance instrumentale par les stimuli (c'est-à-dire le transfert pavlovien-instrumental ) nécessite le noyau accumbens. L'apprentissage instrumental habituel et orienté vers un objectif dépend respectivement du striatum latéral et du striatum médial.
Français Au cours de l'apprentissage instrumental, des changements opposés dans le rapport des récepteurs AMPA aux récepteurs NMDA et ERK phosphorylés se produisent dans les MSN de type D1 et de type D2 qui constituent respectivement les voies directes et indirectes . Ces changements dans la plasticité synaptique et l'apprentissage qui l'accompagne dépendent de l'activation des récepteurs striataux D1 et NMDA. La cascade intracellulaire activée par les récepteurs D1 implique le recrutement de la protéine kinase A et, par la phosphorylation résultante de DARPP-32 , l'inhibition des phosphatases qui désactivent ERK. Les récepteurs NMDA activent ERK par une voie Ras-Raf-MEK-ERK différente mais interdépendante . L'activation d'ERK médiée par le NMDA seul est auto-limitée, car l'activation du NMDA inhibe également l'inhibition médiée par la PKA des phosphatases désactivant ERK. Cependant, lorsque les cascades D1 et NMDA sont co-activées, elles fonctionnent en synergie et l'activation résultante de l'ERK régule la plasticité synaptique sous la forme d'une restructuration de la colonne vertébrale, du transport des récepteurs AMPA, de la régulation de CREB et de l'augmentation de l'excitabilité cellulaire via l'inhibition de Kv4.2 .
Troubles
Dépendance
La surexpression de ΔFosB (DeltaFosB) – un facteur de transcription génétique – dans les neurones épineux moyens de type D1 du noyau accumbens est le facteur commun crucial parmi pratiquement toutes les formes de dépendance (c'est-à-dire les dépendances comportementales et les dépendances aux drogues ) qui induit un comportement et une plasticité neuronale liés à la dépendance . En particulier, ΔFosB favorise l'auto-administration , la sensibilisation à la récompense et les effets de sensibilisation croisée à la récompense parmi des drogues et des comportements addictifs spécifiques. Certaines modifications épigénétiques des queues de protéines d'histones (c'est-à-dire les modifications d'histones) dans des régions spécifiques du cerveau sont également connues pour jouer un rôle crucial dans la base moléculaire des dépendances .
Les drogues et les comportements addictifs sont gratifiants et renforçants (c'est-à-dire addictifs ) en raison de leurs effets sur la voie de récompense de la dopamine .
L' hypothalamus latéral et le faisceau médial du prosencéphale ont été les sites de récompense de stimulation cérébrale les plus fréquemment étudiés, en particulier dans les études sur les effets des drogues sur la récompense de stimulation cérébrale. Le système de neurotransmetteurs qui a été le plus clairement identifié avec les actions de formation d'accoutumance des drogues est le système dopaminergique mésolimbique, avec ses cibles efférentes dans le noyau accumbens et ses afférences GABAergiques locales. Les actions de récompense pertinentes de l'amphétamine et de la cocaïne se situent dans les synapses dopaminergiques du noyau accumbens et peut-être du cortex préfrontal médian . Les rats apprennent également à appuyer sur un levier pour les injections de cocaïne dans le cortex préfrontal médian, ce qui fonctionne en augmentant le renouvellement de la dopamine dans le noyau accumbens. La nicotine infusée directement dans le noyau accumbens augmente également la libération locale de dopamine, vraisemblablement par une action présynaptique sur les terminaisons dopaminergiques de cette région. Les récepteurs nicotiniques se localisent dans les corps cellulaires dopaminergiques et les injections locales de nicotine augmentent la décharge des cellules dopaminergiques qui est essentielle à la récompense nicotinique. Certaines autres drogues créant une dépendance sont également susceptibles de diminuer la production des neurones épineux moyens en conséquence, malgré l'activation des projections dopaminergiques. Pour les opiacés, le site de seuil le plus bas pour les effets de récompense implique des actions sur les neurones GABAergiques dans l' aire tegmentale ventrale , un site secondaire des actions de récompense des opiacés sur les neurones de sortie épineux moyens du noyau accumbens. Ainsi, les éléments suivants constituent le cœur du circuit de récompense médicamenteuse actuellement caractérisé ; Afférents GABAergiques aux neurones dopaminergiques mésolimbiques (substrat principal de la récompense des opiacés), aux neurones dopaminergiques mésolimbiques eux-mêmes (substrat principal de la récompense des stimulants psychomoteurs) et efférents GABAergiques aux neurones dopaminergiques mésolimbiques (un site secondaire de la récompense des opiacés).
Motivation
La saillance motivationnelle dysfonctionnelle apparaît dans un certain nombre de symptômes et de troubles psychiatriques. L'anhédonie , traditionnellement définie comme une capacité réduite à ressentir du plaisir, a été réexaminée comme reflétant une saillance incitative émoussée, car la plupart des populations anhédoniques présentent un « goût » intact. À l'autre extrémité du spectre, une saillance incitative accrue qui se rétrécit pour des stimuli spécifiques est caractéristique des addictions comportementales et aux drogues. Dans le cas de la peur ou de la paranoïa, le dysfonctionnement peut résider dans une saillance aversive élevée . Dans la littérature moderne, l'anhédonie est associée aux deux formes proposées de plaisir, « anticipatoire » et « consommatoire ».
Des études de neuroimagerie sur des diagnostics associés à l'anhédonie ont signalé une activité réduite dans l'OFC et le striatum ventral. Une méta-analyse a signalé que l'anhédonie était associée à une réponse neuronale réduite à l'anticipation de la récompense dans le noyau caudé, le putamen, le noyau accumbens et le cortex préfrontal médian (mPFC).
Troubles de l'humeur
Certains types de dépression sont associés à une motivation réduite, évaluée par la volonté de fournir des efforts pour une récompense. Ces anomalies ont été provisoirement liées à une activité réduite dans les zones du striatum, et bien que les anomalies dopaminergiques soient supposées jouer un rôle, la plupart des études explorant la fonction dopaminergique dans la dépression ont rapporté des résultats contradictoires. Bien que les études post-mortem et de neuroimagerie aient trouvé des anomalies dans de nombreuses régions du système de récompense, peu de résultats sont reproduits de manière cohérente. Certaines études ont rapporté une activité réduite du NAcc, de l'hippocampe, du cortex préfrontal médian (mPFC) et du cortex orbitofrontal (OFC), ainsi qu'une activité élevée de l'amygdale basolatérale et du cortex cingulaire sous-génital (sgACC) pendant les tâches liées à la récompense ou aux stimuli positifs. Ces anomalies de neuroimagerie sont complétées par peu de recherches post-mortem, mais les quelques recherches qui ont été effectuées suggèrent une réduction des synapses excitatrices dans le mPFC. L'activité réduite dans le mPFC pendant les tâches liées à la récompense semble être localisée dans les régions plus dorsales (c'est-à-dire le cortex cingulaire prégénual ), tandis que le sgACC plus ventral est hyperactif dans la dépression.
Les tentatives d'étude des circuits neuronaux sous-jacents dans les modèles animaux ont également donné des résultats contradictoires. Deux paradigmes sont couramment utilisés pour simuler la dépression, la défaite sociale chronique (CSDS) et le stress chronique léger (CMS), bien qu'il en existe plusieurs. Le CSDS produit une préférence réduite pour le saccharose, des interactions sociales réduites et une immobilité accrue dans le test de nage forcée. Le CMS réduit de la même manière la préférence pour le saccharose et le désespoir comportemental tel qu'évalué par les tests de suspension de la queue et de nage forcée. Les animaux sensibles au CSDS présentent une augmentation des décharges phasiques du VTA et l'inhibition des projections VTA-NAcc atténue les déficits comportementaux induits par le CSDS. Cependant, l'inhibition des projections VTA - mPFC exacerbe le retrait social. D'autre part, les réductions associées au CMS de la préférence pour le saccharose et de l'immobilité ont été atténuées et exacerbées par l'excitation et l'inhibition du VTA, respectivement. Bien que ces différences puissent être attribuables à des protocoles de stimulation différents ou à des paradigmes de traduction médiocres, des résultats variables peuvent également résider dans la fonctionnalité hétérogène des régions liées à la récompense.
La stimulation optogénétique du mPFC dans son ensemble produit des effets antidépresseurs. Cet effet semble localisé à l'homologue rongeur du pgACC (le cortex prélimbique), car la stimulation de l'homologue rongeur du sgACC (le cortex infralimbique) ne produit aucun effet comportemental. De plus, la stimulation cérébrale profonde dans le cortex infralimbique, qui est censée avoir un effet inhibiteur, produit également un effet antidépresseur. Cette découverte est cohérente avec l'observation selon laquelle l'inhibition pharmacologique du cortex infralimbique atténue les comportements dépressifs.
Schizophrénie
La schizophrénie est associée à des déficits de motivation, généralement regroupés sous d'autres symptômes négatifs tels qu'une réduction de la parole spontanée . L'expérience de « l'appréciation » est souvent rapportée comme intacte, à la fois sur le plan comportemental et neuronal, bien que les résultats puissent être spécifiques à certains stimuli, tels que les récompenses monétaires. De plus, l'apprentissage implicite et les tâches simples liées à la récompense sont également intacts dans la schizophrénie. Au contraire, les déficits du système de récompense sont apparents lors de tâches liées à la récompense qui sont cognitivement complexes. Ces déficits sont associés à la fois à une activité striatale et OFC anormale, ainsi qu'à des anomalies dans des régions associées à des fonctions cognitives telles que le cortex préfrontal dorsolatéral (DLPFC).
Trouble déficitaire de l'attention avec hyperactivité
Chez les personnes atteintes de TDAH , les aspects fondamentaux du système de récompense sont sous-actifs, ce qui rend difficile la récompense des activités régulières. Les personnes atteintes de ce trouble ressentent un regain de motivation après qu'un comportement à forte stimulation déclenche une libération de dopamine. Après ce regain et cette récompense, le retour aux niveaux de base entraîne une baisse immédiate de la motivation.
Les troubles de la fonction dopaminergique et sérotoninergique sont considérés comme des facteurs clés du TDAH. Ces troubles peuvent conduire à un dysfonctionnement exécutif tel qu'une dysrégulation du traitement de la récompense et un dysfonctionnement motivationnel, notamment l'anhédonie.
Histoire
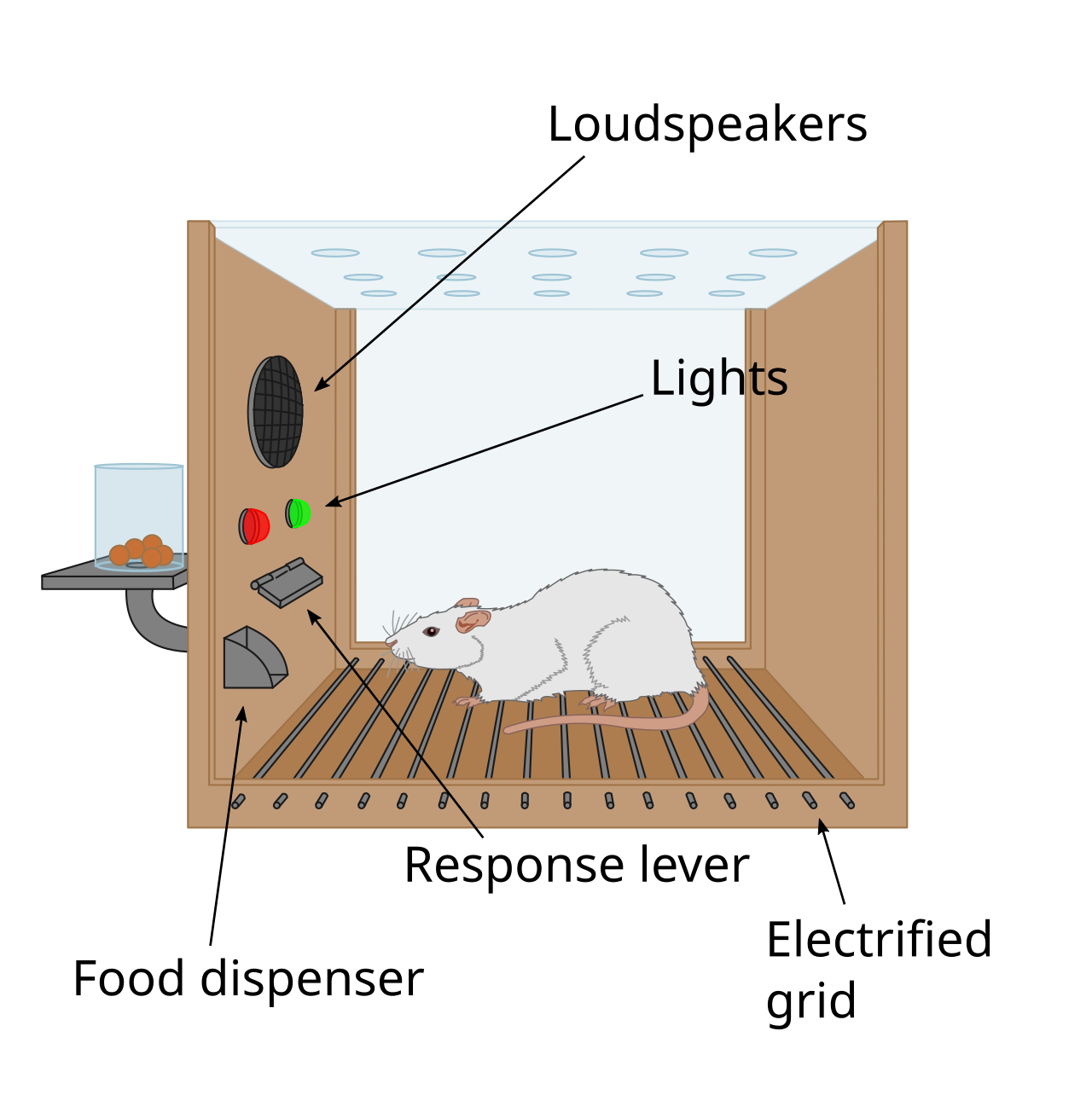
Le premier indice de l’existence d’un système de récompense dans le cerveau est venu avec une découverte accidentelle de James Olds et Peter Milner en 1954. Ils ont découvert que les rats exécutaient des comportements tels que presser une barre, pour administrer une brève décharge de stimulation électrique à des endroits spécifiques de leur cerveau. Ce phénomène est appelé auto-stimulation intracrânienne ou récompense par stimulation cérébrale . En général, les rats appuient sur un levier des centaines ou des milliers de fois par heure pour obtenir cette stimulation cérébrale, s’arrêtant seulement lorsqu’ils sont épuisés. En essayant d’apprendre aux rats comment résoudre des problèmes et courir dans des labyrinthes, la stimulation de certaines régions du cerveau où la stimulation a été trouvée semblait donner du plaisir aux animaux. Ils ont essayé la même chose avec les humains et les résultats ont été similaires. L’explication de la raison pour laquelle les animaux s’engagent dans un comportement qui n’a aucune valeur pour la survie d’eux-mêmes ou de leur espèce est que la stimulation cérébrale active le système sous-jacent à la récompense.
En 1954, les chercheurs James Olds et Peter Milner ont découvert que la stimulation électrique à faible tension de certaines régions du cerveau du rat agissait comme une récompense pour apprendre aux animaux à parcourir des labyrinthes et à résoudre des problèmes. Il semble que la stimulation de ces parties du cerveau ait donné du plaisir aux animaux, et dans des travaux ultérieurs, les humains ont rapporté des sensations agréables résultant d'une telle stimulation. Lorsque des rats ont été testés dans des boîtes Skinner où ils pouvaient stimuler le système de récompense en appuyant sur un levier, les rats ont appuyé pendant des heures. Les recherches menées au cours des deux décennies suivantes ont établi que la dopamine est l'un des principaux produits chimiques facilitant la signalisation neuronale dans ces régions, et la dopamine a été suggérée comme étant le « produit chimique du plaisir » du cerveau.
Ivan Pavlov était un psychologue qui a utilisé le système de récompense pour étudier le conditionnement classique . Pavlov a utilisé le système de récompense en récompensant les chiens avec de la nourriture après qu'ils aient entendu une cloche ou un autre stimulus. Pavlov récompensait les chiens de sorte que les chiens associent la nourriture, la récompense, à la cloche, le stimulus. Edward L. Thorndike a utilisé le système de récompense pour étudier le conditionnement opérant. Il a commencé par mettre des chats dans une boîte à énigmes et en plaçant de la nourriture à l'extérieur de la boîte pour que le chat veuille s'échapper. Les chats ont travaillé pour sortir de la boîte à énigmes pour atteindre la nourriture. Bien que les chats aient mangé la nourriture après s'être échappés de la boîte, Thorndike a appris que les chats tentaient de s'échapper de la boîte sans la récompense de la nourriture. Thorndike a utilisé les récompenses de la nourriture et de la liberté pour stimuler le système de récompense des chats. Thorndike a utilisé cela pour voir comment les chats ont appris à s'échapper de la boîte. Plus récemment, Ivan De Araujo et ses collègues ont utilisé des nutriments à l'intérieur de l'intestin pour stimuler le système de récompense via le nerf vague.
Autres espèces
Les animaux apprennent rapidement à appuyer sur une barre pour obtenir une injection d'opiacés directement dans le tegmentum du mésencéphale ou dans le noyau accumbens . Ces mêmes animaux ne travaillent pas pour obtenir les opiacés si les neurones dopaminergiques de la voie mésolimbique sont inactivés. Dans cette perspective, les animaux, comme les humains, adoptent des comportements qui augmentent la libération de dopamine.
Kent Berridge , chercheur en neurosciences affectives , a découvert que les goûts sucrés ( aimé ) et amer ( détesté ) produisaient des expressions orofaciales distinctes , et ces expressions étaient affichées de la même manière par les nouveau-nés humains, les orangs-outans et les rats. Cela prouve que le plaisir (en particulier le fait d'aimer ) a des caractéristiques objectives et est essentiellement le même chez diverses espèces animales. La plupart des études en neurosciences ont montré que plus la récompense libère de dopamine, plus la récompense est efficace. C'est ce qu'on appelle l'impact hédonique, qui peut être modifié par l'effort fourni pour la récompense et par la récompense elle-même. Berridge a découvert que le blocage des systèmes dopaminergiques ne semblait pas modifier la réaction positive à quelque chose de sucré (mesurée par l'expression faciale). En d'autres termes, l'impact hédonique ne changeait pas en fonction de la quantité de sucre. Cela contredit l'hypothèse conventionnelle selon laquelle la dopamine est un médiateur du plaisir. Même avec des altérations de la dopamine plus intenses, les données semblent rester constantes. Cependant, une étude clinique de janvier 2019 qui a évalué l’effet d’un précurseur de la dopamine ( lévodopa ), d’un antagoniste ( rispéridone ) et d’un placebo sur les réponses de récompense à la musique – y compris le degré de plaisir ressenti pendant les frissons musicaux , tel que mesuré par des changements dans l’activité électrodermique ainsi que des évaluations subjectives – a révélé que la manipulation de la neurotransmission de la dopamine régule de manière bidirectionnelle la cognition du plaisir (en particulier, l’ impact hédonique de la musique ) chez les sujets humains. Cette recherche a démontré qu’une neurotransmission accrue de la dopamine agit comme une condition sine qua non pour des réactions hédoniques agréables à la musique chez les humains.
Berridge a développé l' hypothèse de la saillance incitative pour traiter l' aspect désir des récompenses. Elle explique l'usage compulsif de drogues par les toxicomanes même lorsque la drogue ne produit plus d'euphorie, et les envies ressenties même après que l'individu a terminé son sevrage. Certains toxicomanes répondent à certains stimuli impliquant des changements neuronaux causés par les drogues. Cette sensibilisation dans le cerveau est similaire à l'effet de la dopamine car des réactions de désir et d'appréciation se produisent. Les cerveaux et les comportements humains et animaux connaissent des changements similaires en ce qui concerne les systèmes de récompense car ces systèmes sont si importants.