En génétique et en biochimie , le séquençage consiste à déterminer la structure primaire (parfois appelée à tort séquence primaire) d'un biopolymère non ramifié . Le séquençage donne lieu à une représentation linéaire symbolique appelée séquence qui résume succinctement une grande partie de la structure au niveau atomique de la molécule séquencée.
Séquençage de l'ADN
Le séquençage de l'ADN est le processus de détermination de l' ordre des nucléotides d'un fragment d'ADN donné . Jusqu'à présent, la plupart des séquençages de l'ADN ont été effectués à l'aide de la méthode de terminaison de chaîne développée par Frederick Sanger . Cette technique utilise la terminaison spécifique de la séquence d'une réaction de synthèse de l'ADN à l'aide de substrats nucléotidiques modifiés. Cependant, de nouvelles technologies de séquençage telles que le pyroséquençage gagnent une part croissante du marché du séquençage. Plus de données génomiques sont désormais produites par pyroséquençage que par séquençage de l'ADN de Sanger. Le pyroséquençage a permis un séquençage rapide du génome. Les génomes bactériens peuvent être séquencés en une seule fois avec une couverture à plusieurs reprises grâce à cette technique. Cette technique a également été utilisée pour séquencer le génome de James Watson récemment.
La séquence d'ADN encode les informations nécessaires à la survie et à la reproduction des êtres vivants. Déterminer la séquence est donc utile dans la recherche fondamentale sur les raisons et les modalités de vie des organismes, ainsi que dans les disciplines appliquées. En raison de l'importance clé de l'ADN pour les êtres vivants, la connaissance des séquences d'ADN est utile dans pratiquement tous les domaines de la recherche biologique. Par exemple, en médecine, elle peut être utilisée pour identifier, diagnostiquer et éventuellement développer des traitements pour les maladies génétiques. De même, la recherche sur les agents pathogènes peut conduire à des traitements pour les maladies contagieuses. La biotechnologie est une discipline en plein essor, avec le potentiel de créer de nombreux produits et services utiles.
La courbe de Carlson est un terme inventé par The Economist pour décrire l'équivalent biotechnologique de la loi de Moore , et porte le nom de l'auteur Rob Carlson. Carlson a prédit avec précision que le temps de doublement des technologies de séquençage de l'ADN (mesuré par le coût et les performances) serait au moins aussi rapide que la loi de Moore. Les courbes de Carlson illustrent les diminutions rapides (dans certains cas hyperexponentielles) des coûts et les augmentations des performances d'une variété de technologies, y compris le séquençage de l'ADN, la synthèse de l'ADN et une gamme d'outils physiques et informatiques utilisés dans l'expression des protéines et dans la détermination des structures des protéines.
Séquençage de Sanger
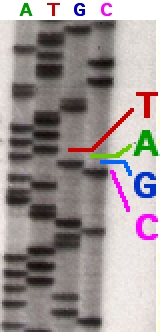
Dans le séquençage de terminateur de chaîne (séquençage de Sanger), l'extension est initiée à un site spécifique sur l'ADN modèle en utilisant une courte « amorce » oligonucléotidique complémentaire à l'ADN modèle dans cette région. L'amorce oligonucléotidique est étendue à l'aide d'une ADN polymérase , une enzyme qui réplique l'ADN. L'amorce et l'ADN polymérase comprennent les quatre bases désoxynucléotidiques (éléments constitutifs de l'ADN), ainsi qu'une faible concentration d'un nucléotide de terminaison de chaîne (le plus souvent un di- désoxynucléotide). Les désoxynucléotides manquent de groupe OH à la fois en position 2' et 3' de la molécule de ribose, donc une fois qu'ils sont insérés dans une molécule d'ADN, ils l'empêchent d'être davantage allongée. Dans ce séquenceur, quatre vaisseaux différents sont utilisés, chacun contenant uniquement les quatre didésoxyribonucléotides ; l'incorporation des nucléotides de terminaison de chaîne par l'ADN polymérase dans une position aléatoire donne lieu à une série de fragments d'ADN apparentés, de différentes tailles, qui se terminent par un didéoxyribonucléotide donné. Les fragments sont ensuite séparés par taille par électrophorèse dans un gel de polyacrylamide en plaque, ou plus communément maintenant, dans un tube en verre étroit (capillaire) rempli d'un polymère visqueux.
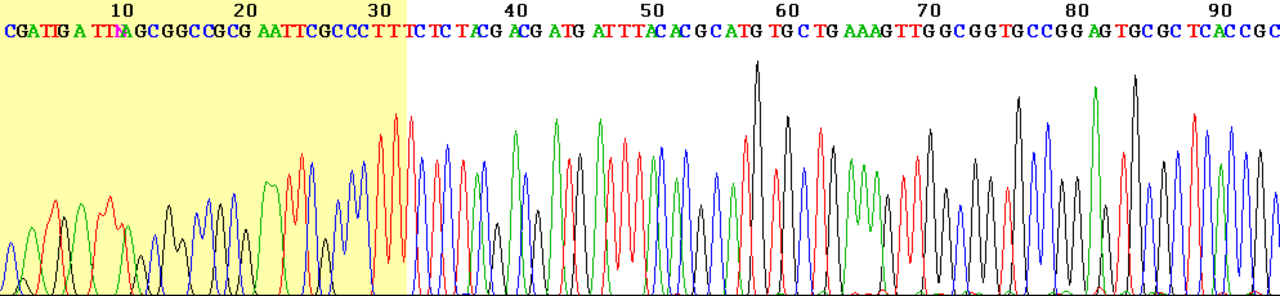
Une alternative au marquage de l'amorce consiste à marquer les terminateurs, ce que l'on appelle communément le « séquençage par terminateur de colorant ». Le principal avantage de cette approche est que l'ensemble du séquençage peut être effectué en une seule réaction, plutôt que les quatre nécessaires avec l'approche par amorce marquée. Cela est accompli en marquant chacun des terminateurs de chaîne didésoxynucléotidique avec un colorant fluorescent distinct, qui émet une fluorescence à une longueur d'onde différente . Cette méthode est plus facile et plus rapide que l'approche par amorce de colorant, mais peut produire des pics de données plus irréguliers (des hauteurs différentes), en raison d'une différence dépendante du modèle dans l'incorporation des grands terminateurs de chaîne de colorant. Ce problème a été considérablement réduit avec l'introduction de nouvelles enzymes et de nouveaux colorants qui minimisent la variabilité d'incorporation. Cette méthode est désormais utilisée pour la grande majorité des réactions de séquençage car elle est à la fois plus simple et moins chère. La principale raison à cela est que les amorces n'ont pas besoin d'être étiquetées séparément (ce qui peut représenter une dépense importante pour une amorce personnalisée à usage unique), bien que ce soit moins un problème avec les amorces « universelles » fréquemment utilisées. Cela évolue rapidement en raison de la rentabilité croissante des systèmes de deuxième et troisième génération d'Illumina, 454, ABI, Helicos et Dover.
Pyroséquençage
La méthode de pyroséquençage est basée sur la détection de la libération de pyrophosphate lors de l'incorporation de nucléotides. Avant de réaliser le pyroséquençage, le brin d'ADN à séquencer doit être amplifié par PCR. Ensuite, l'ordre dans lequel les nucléotides doivent être ajoutés dans le séquenceur est choisi (c'est-à-dire GATC). Lorsqu'un nucléotide spécifique est ajouté, si l'ADN polymérase l'incorpore dans la chaîne en croissance, le pyrophosphate est libéré et converti en ATP par l'ATP sulfurylase. L'ATP alimente l'oxydation de la luciférase par l'intermédiaire de la luciférase ; cette réaction génère un signal lumineux enregistré sous la forme d'un pic de pyrogramme. De cette façon, l'incorporation de nucléotides est corrélée à un signal. Le signal lumineux est proportionnel à la quantité de nucléotides incorporés lors de la synthèse du brin d'ADN (c'est-à-dire que deux nucléotides incorporés correspondent à deux pics de pyrogramme). Lorsque les nucléotides ajoutés ne sont pas incorporés dans la molécule d'ADN, aucun signal n'est enregistré ; l'enzyme apyrase élimine tout nucléotide non incorporé restant dans la réaction. Cette méthode ne nécessite ni nucléotides marqués par fluorescence ni électrophorèse sur gel. Le pyroséquençage, développé par Pål Nyrén et Mostafa Ronaghi DNA, a été commercialisé par Biotage (pour le séquençage à faible débit) et 454 Life Sciences (pour le séquençage à haut débit). Cette dernière plateforme séquence environ 100 mégabases [maintenant jusqu'à 400 mégabases] en sept heures avec une seule machine. Dans la méthode basée sur la matrice (commercialisée par 454 Life Sciences), l'ADN monocaténaire est recuit en billes et amplifié par EmPCR . Ces billes liées à l'ADN sont ensuite placées dans des puits sur une puce à fibre optique avec des enzymes qui produisent de la lumière en présence d' ATP . Lorsque les nucléotides libres sont lavés sur cette puce, de la lumière est produite car l'ATP est généré lorsque les nucléotides se joignent à leurs paires de bases complémentaires . L'ajout d'un (ou plusieurs) nucléotide(s) entraîne une réaction qui génère un signal lumineux enregistré par la caméra CCD de l'instrument. L'intensité du signal est proportionnelle au nombre de nucléotides, par exemple des segments d'homopolymères, incorporés dans un seul flux de nucléotides. [1]
Véritable séquençage d'une seule molécule
Séquençage à grande échelle
Bien que les méthodes ci-dessus décrivent diverses méthodes de séquençage, des termes connexes distincts sont utilisés lorsqu'une grande partie d'un génome est séquencée. Plusieurs plates-formes ont été développées pour effectuer le séquençage de l'exome (un sous-ensemble de l'ADN de tous les chromosomes qui codent les gènes) ou le séquençage du génome entier (séquençage de l'ADN nucléaire d'un être humain).
Séquençage de l'ARN
L'ARN est moins stable dans la cellule et plus sujet à l'attaque expérimentale par les nucléases. Comme l'ARN est généré par transcription à partir de l'ADN, l'information est déjà présente dans l'ADN de la cellule. Cependant, il est parfois souhaitable de séquencer des molécules d'ARN . Alors que le séquençage de l'ADN donne un profil génétique d'un organisme, le séquençage de l'ARN reflète uniquement les séquences qui sont activement exprimées dans les cellules. Pour séquencer l'ARN, la méthode habituelle consiste d'abord à inverser la transcription de l'ARN extrait de l'échantillon pour générer des fragments d'ADNc. Ceux-ci peuvent ensuite être séquencés comme décrit ci-dessus. La majeure partie de l'ARN exprimé dans les cellules est constituée d'ARN ribosomiques ou de petits ARN , nuisibles à la traduction cellulaire, mais qui ne sont souvent pas l'objet d'une étude. Cette fraction peut cependant être éliminée in vitro pour enrichir l'ARN messager, également inclus, qui présente généralement un intérêt. Dérivés des exons, ces ARNm doivent être ultérieurement traduits en protéines qui soutiennent des fonctions cellulaires particulières. Le profil d'expression indique donc l'activité cellulaire, particulièrement recherchée dans les études de maladies, de comportement cellulaire, de réponses à des réactifs ou à des stimuli. Les molécules d'ARN eucaryotes ne sont pas nécessairement colinéaires avec leur matrice d'ADN, car les introns sont excisés. Cela confère une certaine complexité pour cartographier les séquences lues jusqu'au génome et ainsi identifier leur origine. Pour plus d'informations sur les capacités du séquençage de nouvelle génération appliqué aux transcriptomes entiers, voir : RNA-Seq et MicroRNA Sequencing .
Séquençage des protéines
Les méthodes permettant de réaliser le séquençage des protéines comprennent :
- Dégradation d'Edman
- Empreintes digitales de masse peptidique
- Spectrométrie de masse
- Digestifs de protéase
Si le gène codant la protéine est connu, il est actuellement beaucoup plus facile de séquencer l'ADN et d'en déduire la séquence protéique. La détermination d'une partie de la séquence d'acides aminés d'une protéine (souvent une extrémité) par l'une des méthodes ci-dessus peut suffire à identifier un clone porteur de ce gène.
Séquençage des polysaccharides
Bien que les polysaccharides soient également des biopolymères, il n'est pas si courant de parler de « séquençage » d'un polysaccharide, pour plusieurs raisons. Bien que de nombreux polysaccharides soient linéaires, beaucoup ont des ramifications. De nombreuses unités différentes ( monosaccharides individuels ) peuvent être utilisées et liées de différentes manières. Cependant, la principale raison théorique est que, alors que les autres polymères répertoriés ici sont principalement générés de manière « dépendante du modèle » par une enzyme processive, chaque jonction individuelle dans un polysaccharide peut être formée par une enzyme différente . Dans de nombreux cas, l'assemblage n'est pas spécifié de manière unique ; selon l'enzyme qui agit, une des différentes unités peut être incorporée. Cela peut conduire à la formation d'une famille de molécules similaires. Cela est particulièrement vrai pour les polysaccharides végétaux. Les méthodes de détermination de la structure des oligosaccharides et des polysaccharides comprennent la spectroscopie RMN et l'analyse de la méthylation.