Les collemboles (classe Collembola ) constituent le plus important des trois groupes d' hexapodes modernes qui ne sont plus considérés comme des insectes , les deux autres étant les Protoures et les Diploures . Bien que ces trois groupes soient parfois regroupés dans la classe des Entognathes en raison de leurs pièces buccales internes , ils ne semblent pas plus proches les uns des autres que des insectes, qui possèdent des pièces buccales externes. On compte plus de 9 000 espèces.
Les collemboles sont des organismes omnivores et libres qui préfèrent les milieux humides. Ils ne participent pas directement à la décomposition de la matière organique, mais y contribuent indirectement par sa fragmentation et la régulation des communautés microbiennes du sol. Le mot Collembola vient du grec ancien signifiant « colle », et signifiant « cheville » ; ce nom leur a été donné en raison de l'existence du collophore , que l'on pensait auparavant leur permettre d'adhérer aux surfaces et de se stabiliser.
Les premières études de séquences d'ADN suggéraient que les collemboles représentaient une lignée évolutive distincte des autres hexapodes , mais d'autres études contestent cette hypothèse ; cela semble dû à des schémas d' évolution moléculaire très divergents chez les arthropodes . Les ajustements apportés à la classification taxonomique traditionnelle des collemboles reflètent l'incompatibilité occasionnelle des regroupements traditionnels avec la cladistique moderne : lorsqu'ils étaient inclus parmi les insectes , ils étaient classés comme un ordre ; au sein des Entognathes , ils sont classés comme une sous-classe . S'ils sont considérés comme faisant partie des hexapodes , ils sont élevés au rang de classe à part entière .
Morphologie


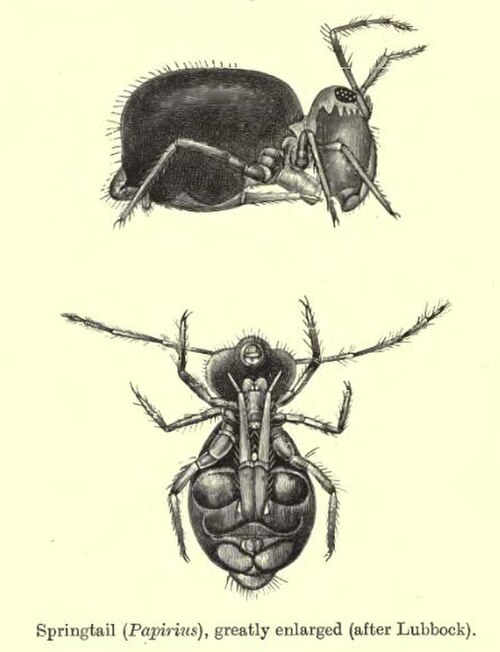
Les collemboles mesurent généralement moins de de long, possèdent six segments abdominaux ou moins et un appendice tubulaire (le collophore ou tube ventral) muni de vésicules collantes réversibles , qui fait saillie ventralement du premier segment abdominal. On pense qu'il est impliqué dans l'absorption et l'équilibre des fluides, l'excrétion et l'orientation de l'organisme. La plupart des espèces possèdent un appendice abdominal en forme de queue appelé furcula (ou furca). Situé sur la face ventrale du quatrième segment abdominal, il est replié sous le corps et maintenu sous tension par une petite structure appelée rétinaculum (ou tenaculum). Lorsqu'il est relâché, il claque contre le substrat, propulsant le collembole dans les airs et lui permettant de se déplacer et de fuir rapidement. Tout cela se déroule en seulement 18 millisecondes.
Les collemboles ont également la capacité de réduire leur taille corporelle jusqu'à 30 % par mues successives si les températures augmentent suffisamment. Cette réduction est génétiquement contrôlée. Étant donné que la hausse des températures accroît le métabolisme et les besoins énergétiques des organismes, la réduction de la taille corporelle est avantageuse pour leur survie.
Les Poduromorpha et les Entomobryomorpha possèdent un corps allongé, tandis que les Symphypleona et les Neelipleona ont un corps globulaire. Les Collemboles sont dépourvus de système respiratoire trachéal , ce qui les oblige à respirer à travers une cuticule poreuse , à l'exception des deux familles Sminthuridae et Actaletidae , qui présentent une unique paire de stigmates entre la tête et le thorax , formant ainsi un système trachéal rudimentaire, bien que pleinement fonctionnel. La variance anatomique (variabilité de la forme de vie) présente entre les différentes espèces dépend en partie de leur distribution verticale à travers les différentes strates des écosystèmes terrestres . Les espèces vivant en surface sont généralement plus grandes, ont des pigments plus foncés, des antennes plus longues et une furcula fonctionnelle. Les espèces vivant en subsurface sont généralement non pigmentées, ont un corps allongé et une furcula réduite. On peut les classer en quatre catégories principales selon leur répartition verticale : atmobiotiques, épédaphiques, hémiédaphiques et euédaphiques. Les espèces atmobiotiques vivent sur les macrophytes et à la surface de la litière. Elles mesurent généralement de 8 à 10 millimètres (environ ⅓ de pouce) de long, sont pigmentées, possèdent de longs limbes et un ensemble complet d’ ocelles (photorécepteurs). Les espèces épédaphiques vivent dans les couches supérieures de la litière et sur les troncs d’arbres tombés. Elles sont légèrement plus petites et présentent une pigmentation moins prononcée, ainsi que des limbes et des ocelles moins développés que les espèces atmobiotiques. Les espèces hémiédaphiques vivent dans les couches inférieures de la litière, composées de matière organique en décomposition. Elles mesurent de 1 à 2 millimètres (environ 1/16 de pouce) de long, ont une pigmentation dispersée, des limbes raccourcis et un nombre réduit d’ocelles. Les espèces euédaphiques vivent dans les couches minérales supérieures appelées horizon humifère (ou humipède ). Elles sont plus petites que les espèces hémiédaphiques. Ils possèdent un corps mou et allongé ; ils sont dépourvus de pigmentation et d’ocelles ; et leur furca est réduite ou absente.
Les Poduromorphes se caractérisent par leurs corps allongés et leur segmentation marquée : ils possèdent trois segments thoraciques, six segments abdominaux, dont un prothorax bien développé avec des soies tergales , tandis que le premier segment thoracique des Entomobryomorpha est clairement réduit et ne porte pas de soies.
Le tube digestif des collemboles comprend trois parties principales : l’intestin antérieur , l’intestin moyen et l’intestin postérieur . L’intestin moyen est entouré d’un réseau musculaire et tapissé d’une monocouche de cellules cylindriques ou cuboïdes . Sa fonction est de mélanger et de transporter les aliments de la lumière intestinale vers l’intestin postérieur par contraction. De nombreuses espèces de bactéries , d’archées et de champignons syntrophes sont présentes dans la lumière intestinale. Ces différentes régions digestives présentent des pH variables, permettant des activités enzymatiques et des populations microbiennes spécifiques. La partie antérieure de l’intestin moyen et de l’intestin postérieur est légèrement acide (pH d’environ 6,0), tandis que la partie postérieure de l’intestin moyen est légèrement alcaline (pH d’environ 8,0). Entre l’intestin moyen et l’intestin postérieur se trouve le pylore , un canal digestif constitué d’un sphincter musculaire . Les tubes de Malpighi sont absents.
Génomique
Du fait de leur petite taille, les collemboles ont été négligés en matière d' analyse génomique . Ils font partie des rares groupes d'arthropodes ne disposant pas de génomes de référence de haute qualité . Bien que certaines séquences génomiques aient été produites par le passé, elles ne répondent pas aux normes actuelles (contig N50 > 1 Mb). L'un des premiers génomes, celui d' Orchesella flavescens , a été présenté en 2025. D'une taille de 270 Mb, il a été assemblé en six échafaudages chromosomiques
Systématique et évolution
Traditionnellement, les collemboles étaient classés en trois ordres : les Arthropleona , les Symphypleona et, plus rarement, les Neelipleona . Les Arthropleona étaient eux-mêmes divisés en deux superfamilles : les Entomobryoidea et les Poduroidea . Cependant, des études phylogénétiques récentes montrent que les Arthropleona sont paraphylétiques . Par conséquent, les Arthropleona sont supprimés des classifications modernes et leurs superfamilles sont reclassées en conséquence, devenant ainsi les ordres Entomobryomorpha et Poduromorpha . Techniquement, les Arthropleona sont donc un synonyme partiel plus récent des Collembola
Le terme Neopleona est essentiellement synonyme de Symphypleona + Neelipleona . Les Neelipleona étaient initialement considérés comme une lignée particulièrement évoluée de Symphypleona, en raison de leur morphologie corporelle globale commune. Cependant, cette morphologie est en réalité totalement différente chez les Neelipleona. Par la suite, on a considéré que les Neelipleona dérivaient des Entomobryomorpha. L'analyse des séquences d'ARNr 18S et 28S suggère néanmoins qu'ils constituent la lignée la plus ancienne des collemboles, ce qui expliquerait leurs apomorphies particulières . Cette relation phylogénétique a également été confirmée par une phylogénie basée sur l'ADNmt et sur des données de génomes entiers .
La phylogénie du génome entier la plus récente soutient quatre ordres de Collemboles :