Domaine des organismes Archées ( / ɑːr ˈ k jeː ə / ⓘ ar- KEE -ə ) est un domaine d' organismes . Traditionnellement, les archées n'incluaient que ses procaryotes , mais ce sens ...
Worldlex WikiFrancaisLecture gratuite
Domaine des organismes
Archées ( / ɑːr ˈ k jeː ə /ⓘ ar-KEE-ə) est undomained'organismes. Traditionnellement, les archées n'incluaient que sesprocaryotes, mais ce sens s'est avéré êtreparaphylétique, carles eucaryotesont évolué à partir des archées. Même si le domaine Archaea inclut les eucaryotes, le terme « archées » (sg.: archéon/ ɑːr ˈ k iː ɒ n /ar-KEE-on, du grec « ἀρχαῖον », qui signifie ancien) en anglais fait toujours généralement référence spécifiquement aux membres procaryotes des archées. Les archées étaient initialementclasséescommedes bactéries, recevant le nom d'archaebactéries (/ ˌ ɑːr k i b æ k ˈ t ɪər i ə /, dans lerègne), mais ce terme est tombé en désuétude.
Les cellules archéennes ont des propriétés uniques qui les distinguent des bactéries et des eucaryotes . Les archées sont elles-mêmes divisées en plusieurs phylums reconnus . La classification est difficile car la plupart n'ont pas été isolées en laboratoire et n'ont été détectées que par leurs séquences génétiques dans des échantillons environnementaux. On ne sait pas si elles sont capables de produire des endospores .
Les archées et les bactéries sont généralement de taille et de forme similaires, bien que quelques archées aient des formes très différentes, comme les cellules plates et carrées de Haloquadratum walsbyi . Malgré cette similitude morphologique avec les bactéries, les archées possèdent des gènes et plusieurs voies métaboliques qui sont plus étroitement liés à ceux des eucaryotes, notamment pour les enzymes impliquées dans la transcription et la traduction . D'autres aspects de la biochimie des archées sont uniques, comme leur dépendance aux lipides éther dans leurs membranes cellulaires , y compris les archéols . Les archées utilisent des sources d'énergie plus diverses que les eucaryotes, allant des composés organiques tels que les sucres, à l'ammoniac , aux ions métalliques ou même à l'hydrogène gazeux . Les Haloarchaea tolérantes au sel utilisent la lumière du soleil comme source d'énergie, et d'autres espèces d'archées fixent le carbone (autotrophie), mais contrairement aux plantes et aux cyanobactéries , aucune espèce connue d'archées ne fait les deux. Les archées se reproduisent de manière asexuée par fission binaire , fragmentation ou bourgeonnement ; contrairement aux bactéries, aucune espèce connue d'archées ne forme d'endospores . Les premières archées observées étaient des extrémophiles , vivant dans des environnements extrêmes tels que des sources chaudes et des lacs salés sans aucun autre organisme. Des outils de détection moléculaire améliorés ont conduit à la découverte d'archées dans presque tous les habitats , y compris le sol, les océans et les marais . Les archées sont particulièrement nombreuses dans les océans, et les archées du plancton pourraient être l'un des groupes d'organismes les plus abondants de la planète.
Les archées jouent un rôle majeur dans la vie sur Terre . Elles font partie du microbiote de tous les organismes. Dans le microbiome humain , elles sont importantes dans l' intestin , la bouche et sur la peau. Leur diversité morphologique, métabolique et géographique leur permet de jouer de multiples rôles écologiques : fixation du carbone, cycle de l'azote , renouvellement des composés organiques et maintien des communautés microbiennes symbiotiques et syntrophiques , par exemple.
Pendant une grande partie du XXe siècle, les procaryotes étaient considérés comme un seul groupe d'organismes et classés en fonction de leur biochimie , de leur morphologie et de leur métabolisme . Les microbiologistes ont essayé de classer les micro-organismes en fonction de la structure de leurs parois cellulaires , de leur forme et des substances qu'ils consomment. En 1965, Emile Zuckerkandl et Linus Pauling ont plutôt proposé d'utiliser les séquences des gènes de différents procaryotes pour déterminer comment ils sont liés les uns aux autres. Cette approche phylogénétique est la principale méthode utilisée aujourd'hui.
Les archées ont été classées pour la première fois séparément des bactéries en 1977 par Carl Woese et George E. Fox , sur la base de leurs gènes d'ARN ribosomique (ARNr). (A cette époque, seuls les méthanogènes étaient connus). Ils ont appelé ces groupes les royaumes des archéobactéries et des eubactéries, bien que d'autres chercheurs les aient traités comme des royaumes ou des sous-royaumes. Woese et Fox ont fourni la première preuve que les archéobactéries constituaient une « lignée de descendance » distincte : 1. l'absence de peptidoglycane dans leurs parois cellulaires, 2. deux coenzymes inhabituels, 3. les résultats du séquençage du gène de l'ARN ribosomique 16S . Pour souligner cette différence, Woese, Otto Kandler et Mark Wheelis ont ensuite proposé de reclasser les organismes en trois domaines naturels connus sous le nom de système à trois domaines : les eucaryotes , les bactéries et les archées, dans ce qui est maintenant connu sous le nom de révolution woesienne .
Le mot archée vient du grec ancien ἀρχαῖα , qui signifie « choses anciennes », car les premiers représentants du domaine Archaea étaient des méthanogènes et on supposait que leur métabolisme reflétait l'atmosphère primitive de la Terre et l'ancienneté des organismes, mais à mesure que de nouveaux habitats étaient étudiés, davantage d'organismes ont été découverts. Des microbes halophiles extrêmes et hyperthermophiles étaient également inclus dans Archaea. Pendant longtemps, les archées ont été considérées comme des extrémophiles qui n'existent que dans des habitats extrêmes tels que les sources chaudes et les lacs salés , mais à la fin du 20e siècle, des archées ont également été identifiées dans des environnements non extrêmes. Aujourd'hui, on sait qu'elles constituent un groupe vaste et diversifié d'organismes abondamment répartis dans la nature. Cette nouvelle appréciation de l'importance et de l'ubiquité des archées est venue de l'utilisation de la réaction en chaîne par polymérase (PCR) pour détecter les procaryotes à partir d'échantillons environnementaux (tels que l'eau ou le sol) en multipliant leurs gènes ribosomiques. Cela permet la détection et l'identification d'organismes qui n'ont pas été cultivés en laboratoire.
La classification des archées, et des procaryotes en général, est un domaine en évolution rapide et controversé. Les systèmes de classification actuels visent à organiser les archées en groupes d'organismes qui partagent des caractéristiques structurelles et des ancêtres communs. Ces classifications s'appuient largement sur l'utilisation de la séquence des gènes d'ARN ribosomique pour révéler les relations entre les organismes ( phylogénétique moléculaire ). La plupart des espèces d'archées cultivables et bien étudiées sont membres de deux embranchements principaux , les « Euryarchaeota » et les Thermoproteota (anciennement Crenarchaeota). D'autres groupes ont été provisoirement créés, comme l'espèce particulière Nanoarchaeum equitans — découverte en 2003 et à laquelle a été attribué son propre embranchement, les « Nanoarchaeota ». Un nouveau embranchement « Korarchaeota » a également été proposé, contenant un petit groupe d'espèces thermophiles inhabituelles partageant des caractéristiques des deux embranchements principaux, mais plus étroitement liées aux Thermoproteota. D'autres espèces d'archées détectées ne sont que vaguement apparentées à l'un de ces groupes, comme les nanoorganismes acidophiles archaïques de la mine Richmond (ARMAN, comprenant Micrarchaeota et Parvarchaeota), qui ont été découverts en 2006 et sont parmi les plus petits organismes connus.
Un superphylum – TACK – qui comprend les Thaumarchaeota (maintenant Nitrososphaerota ), « Aigarchaeota », Crenarchaeota (maintenant Thermoproteota ) et « Korarchaeota » a été proposé en 2011 comme étant lié à l'origine des eucaryotes. Asgard nouvellement découvert et nouvellement nommé a été proposé comme étant plus étroitement lié à l'eucaryote d'origine et un groupe frère de TACK.
En 2013, le superphylum DPANN a été proposé pour regrouper les « Nanoarchaeota », les « Nanohaloarchaeota », les nanoorganismes acidophiles archaïques de Richmond Mine (ARMAN, comprenant les « Micrarchaeota » et les « Parvarchaeota ») et d'autres archées similaires. Ce superphylum archéen englobe au moins 10 lignées différentes et comprend des organismes aux tailles de cellules et de génomes extrêmement petites et aux capacités métaboliques limitées. Par conséquent, le DPANN peut inclure des membres obligatoirement dépendants des interactions symbiotiques, et peut même inclure de nouveaux parasites. Cependant, d'autres analyses phylogénétiques ont révélé que le DPANN ne forme pas un groupe monophylétique et que le regroupement apparent est causé par l'attraction des longues branches (LBA), suggérant que toutes ces lignées appartiennent aux « Euryarchaeota ».
Cladogramme
Selon Tom A. Williams et al. 2017, Castelle & Banfield (2018) et GTDB version 09-RS220 (24 avril 2024) :
La classification des archées en espèces est également controversée. La définition d' espèce d' Ernst Mayr — un groupe d'organismes reproducteurs isolés — ne s'applique pas, car les archées se reproduisent uniquement de manière asexuée.
Les archées présentent des niveaux élevés de transfert horizontal de gènes entre les lignées. Certains chercheurs suggèrent que les individus peuvent être regroupés en populations semblables à celles des espèces, étant donné les génomes très similaires et les transferts de gènes peu fréquents vers/depuis des cellules ayant des génomes moins apparentés, comme dans le genre Ferroplasma . D'un autre côté, des études sur Halorubrum ont révélé un transfert génétique significatif vers/depuis des populations moins apparentées, ce qui limite l'applicabilité du critère. Certains chercheurs se demandent si de telles désignations d'espèces ont une signification pratique.
Les connaissances actuelles sur la diversité génétique des archées sont fragmentaires, de sorte que le nombre total d'espèces ne peut être estimé avec précision. Les estimations du nombre de phylums varient de 18 à 23, dont seulement 8 ont des représentants qui ont été cultivés et étudiés directement. Beaucoup de ces groupes hypothétiques sont connus à partir d'une seule séquence d'ARNr, de sorte que le niveau de diversité reste obscur. Cette situation est également observée chez les bactéries ; de nombreux microbes non cultivés présentent des problèmes similaires de caractérisation.
Embranchements procaryotes
Embranchements valides
Les phylums suivants ont été valablement publiés selon le Code bactériologique :
Les phylums suivants ont été proposés, mais n'ont pas été publiés de manière valide selon le Code bactériologique (y compris ceux qui ont le statut de candidatus ) :
Bien que les fossiles probables de cellules procaryotes datent de près de 3,5 milliards d'années , la plupart des procaryotes n'ont pas de morphologies distinctives et les formes fossiles ne peuvent pas être utilisées pour les identifier comme archées. Au lieu de cela, les fossiles chimiques de lipides uniques sont plus informatifs car de tels composés ne sont pas présents dans d'autres organismes. Certaines publications suggèrent que des restes lipidiques archaïques ou eucaryotes sont présents dans des schistes datant d'il y a 2,7 milliards d'années, bien que ces données aient depuis été remises en question. Ces lipides ont également été détectés dans des roches encore plus anciennes de l'ouest du Groenland . Les plus anciennes traces de ce type proviennent du district d'Isua , qui comprend les plus anciens sédiments connus de la Terre, formés il y a 3,8 milliards d'années. La lignée archéenne pourrait être la plus ancienne qui existe sur Terre.
Woese a soutenu que les bactéries, les archées et les eucaryotes représentent des lignées distinctes de descendance qui ont divergé très tôt d'une colonie ancestrale d'organismes. Une possibilité est que cela se soit produit avant l' évolution des cellules , lorsque l'absence d'une membrane cellulaire typique a permis un transfert latéral de gènes sans restriction , et que les ancêtres communs des trois domaines soient apparus par fixation de sous-ensembles spécifiques de gènes. Il est possible que le dernier ancêtre commun des bactéries et des archées ait été un thermophile , ce qui soulève la possibilité que les températures plus basses soient des « environnements extrêmes » pour les archées, et que les organismes qui vivent dans des environnements plus frais ne soient apparus que plus tard. Étant donné que les archées et les bactéries ne sont pas plus apparentées entre elles qu'elles ne le sont aux eucaryotes, le terme procaryote peut suggérer une fausse similitude entre elles. Cependant, les similitudes structurelles et fonctionnelles entre les lignées se produisent souvent en raison de traits ancestraux partagés ou d' une convergence évolutive . Ces similitudes sont connues sous le nom de grade , et les procaryotes sont mieux considérés comme un grade de vie, caractérisé par des caractéristiques telles que l'absence d'organites liés à une membrane.
Comparaison avec d'autres domaines
Le tableau suivant compare certaines caractéristiques majeures des trois domaines, pour illustrer leurs similitudes et leurs différences.
Les archées ont été séparées en un troisième domaine en raison des grandes différences dans la structure de leur ARN ribosomique. La molécule particulière 16S rRNA est essentielle à la production de protéines dans tous les organismes. Comme cette fonction est si essentielle à la vie, les organismes présentant des mutations dans leur 16S rRNA ont peu de chances de survivre, ce qui conduit à une grande stabilité (mais pas absolue) de la structure de ce polynucléotide au fil des générations. L'ARNr 16S est suffisamment grand pour montrer des variations spécifiques à l'organisme, mais suffisamment petit pour être comparé rapidement. En 1977, Carl Woese, un microbiologiste étudiant les séquences génétiques des organismes, a développé une nouvelle méthode de comparaison qui consistait à diviser l'ARN en fragments qui pouvaient être triés et comparés à d'autres fragments d'autres organismes. Plus les modèles entre les espèces sont similaires, plus elles sont étroitement liées.
Woese a utilisé sa nouvelle méthode de comparaison d'ARNr pour classer et comparer différents organismes. Il a comparé une variété d'espèces et est tombé sur un groupe de méthanogènes avec un ARNr très différent de tous les procaryotes ou eucaryotes connus. Ces méthanogènes étaient beaucoup plus similaires entre eux qu'à d'autres organismes, ce qui a conduit Woese à proposer le nouveau domaine des archées. Ses expériences ont montré que les archées étaient génétiquement plus semblables aux eucaryotes qu'aux procaryotes, même si elles étaient plus semblables aux procaryotes dans leur structure. Cela a conduit à la conclusion que les archées et les eucaryotes partageaient un ancêtre commun plus récent que les eucaryotes et les bactéries. Le développement du noyau s'est produit après la séparation entre les bactéries et cet ancêtre commun.
Une propriété unique aux archées est l'utilisation abondante de lipides liés à l'éther dans leurs membranes cellulaires. Les liaisons éther sont chimiquement plus stables que les liaisons ester trouvées dans les bactéries et les eucaryotes, ce qui peut être un facteur contribuant à la capacité de nombreuses archées à survivre dans des environnements extrêmes qui exercent un stress important sur les membranes cellulaires, comme la chaleur et la salinité extrêmes . L'analyse comparative des génomes archéens a également identifié plusieurs indels de signature moléculaire conservés et des protéines de signature présentes de manière unique dans toutes les archées ou dans différents groupes principaux au sein des archées. Une autre caractéristique unique des archées, trouvée dans aucun autre organisme, est la méthanogenèse (la production métabolique de méthane). Les archées méthanogènes jouent un rôle central dans les écosystèmes avec des organismes qui tirent leur énergie de l'oxydation du méthane, dont beaucoup sont des bactéries, car elles sont souvent une source majeure de méthane dans de tels environnements et peuvent jouer un rôle de producteurs primaires. Les méthanogènes jouent également un rôle essentiel dans le cycle du carbone , en décomposant le carbone organique en méthane, qui est également un gaz à effet de serre majeur.
Cette différence dans la structure biochimique des bactéries et des archées a été expliquée par les chercheurs par des processus évolutifs. On suppose que les deux domaines sont originaires des sources hydrothermales alcalines des grands fonds marins . Au moins deux fois, les microbes ont développé la biosynthèse des lipides et la biochimie des parois cellulaires. Il a été suggéré que le dernier ancêtre commun universel était un organisme non libre. Il aurait pu avoir une membrane perméable composée d'amphiphiles bactériens à chaîne simple (acides gras), y compris des amphiphiles archaïques à chaîne simple (isoprénoïdes). Ceux-ci stabilisent les membranes d'acides gras dans l'eau de mer ; cette propriété pourrait avoir entraîné la divergence des membranes bactériennes et archaïques, « avec la biosynthèse ultérieure des phospholipides donnant naissance aux groupes de tête G1P et G3P uniques des archées et des bactéries respectivement. Si tel est le cas, les propriétés conférées par les isoprénoïdes membranaires placent la division lipidique dès l'origine de la vie ».
Relation avec les bactéries
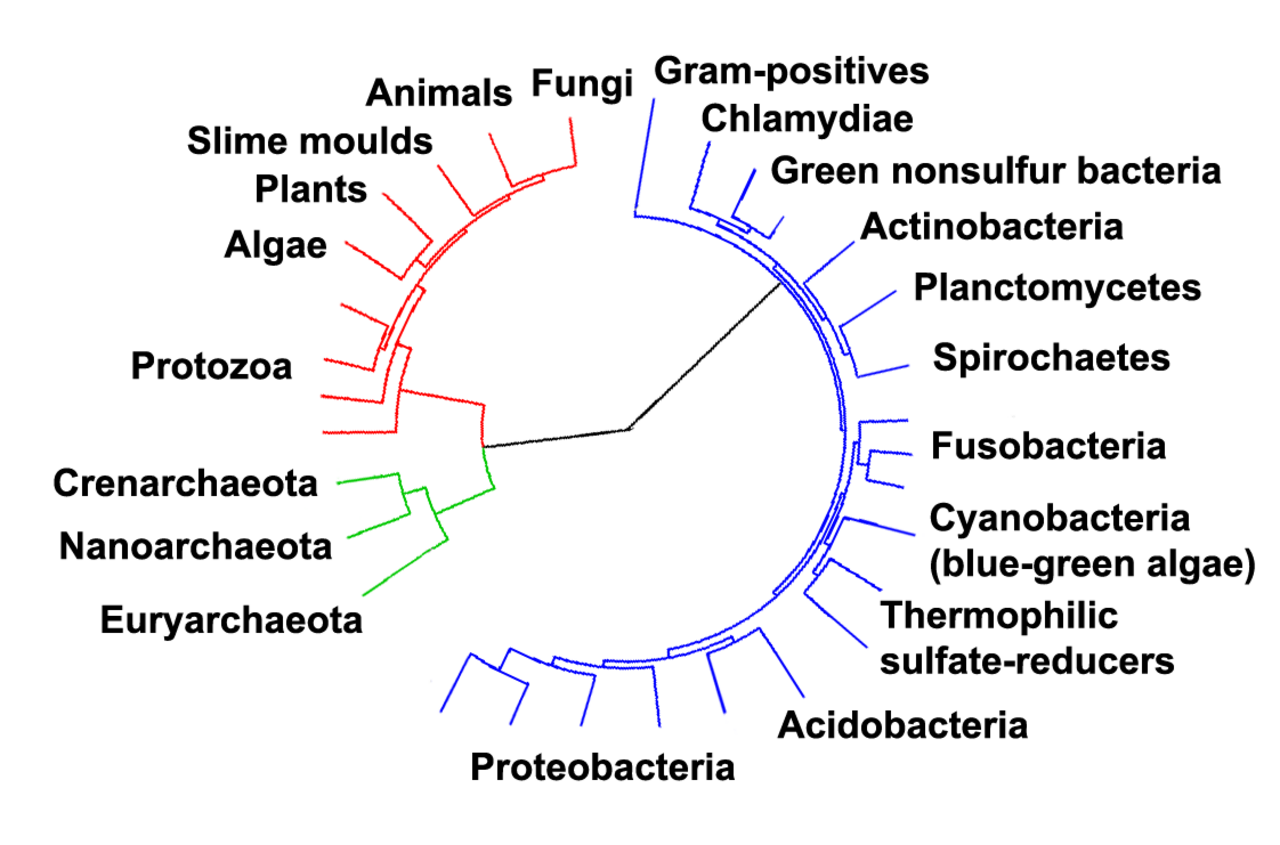
Arbre phylogénétique montrant la relation entre les archées et les autres domaines de la vie. Les eucaryotes sont colorés en rouge, les archées en vert et les bactéries en bleu. Adapté de Ciccarelli et al. (2006)
Les relations entre les trois domaines sont d'une importance capitale pour comprendre l'origine de la vie. La plupart des voies métaboliques , qui sont l'objet de la majorité des gènes d'un organisme, sont communes entre les archées et les bactéries, tandis que la plupart des gènes impliqués dans l'expression du génome sont communs entre les archées et les eucaryotes. Chez les procaryotes, la structure cellulaire des archées est très similaire à celle des bactéries à Gram positif , en grande partie parce que les deux ont une seule bicouche lipidique et contiennent généralement un sacculus épais (exosquelette) de composition chimique variable. Dans certains arbres phylogénétiques basés sur différentes séquences de gènes/protéines d'homologues procaryotes, les homologues archées sont plus étroitement liés à ceux des bactéries à Gram positif. Les archées et les bactéries à Gram positif partagent également des indels conservés dans un certain nombre de protéines importantes, telles que Hsp70 et la glutamine synthétase I ; mais la phylogénie de ces gènes a été interprétée comme révélant un transfert de gènes interdomaines, et pourrait ne pas refléter la ou les relations entre organismes.
Il a été proposé que les archées aient évolué à partir de bactéries Gram-positives en réponse à la pression de sélection des antibiotiques . Ceci est suggéré par l'observation que les archées sont résistantes à une grande variété d'antibiotiques qui sont produits principalement par des bactéries Gram-positives, et que ces antibiotiques agissent principalement sur les gènes qui distinguent les archées des bactéries. L'hypothèse est que la pression sélective vers la résistance générée par les antibiotiques Gram-positifs a finalement été suffisante pour provoquer des changements importants dans de nombreux gènes cibles des antibiotiques, et que ces souches représentaient les ancêtres communs des archées actuelles. L'évolution des archées en réponse à la sélection des antibiotiques, ou à toute autre pression sélective compétitive, pourrait également expliquer leur adaptation à des environnements extrêmes (tels que des températures élevées ou une acidité) comme résultat d'une recherche de niches inoccupées pour échapper aux organismes producteurs d'antibiotiques ; Cavalier-Smith a fait une suggestion similaire, l' hypothèse de Neomura . Cette proposition est également soutenue par d’autres travaux examinant les relations structurelles des protéines et des études suggérant que les bactéries à Gram positif pourraient constituer les premières lignées ramifiées au sein des procaryotes.
La relation évolutive entre les archées et les eucaryotes reste floue. Outre les similitudes dans la structure et la fonction cellulaires qui sont abordées ci-dessous, de nombreux arbres génétiques regroupent les deux.
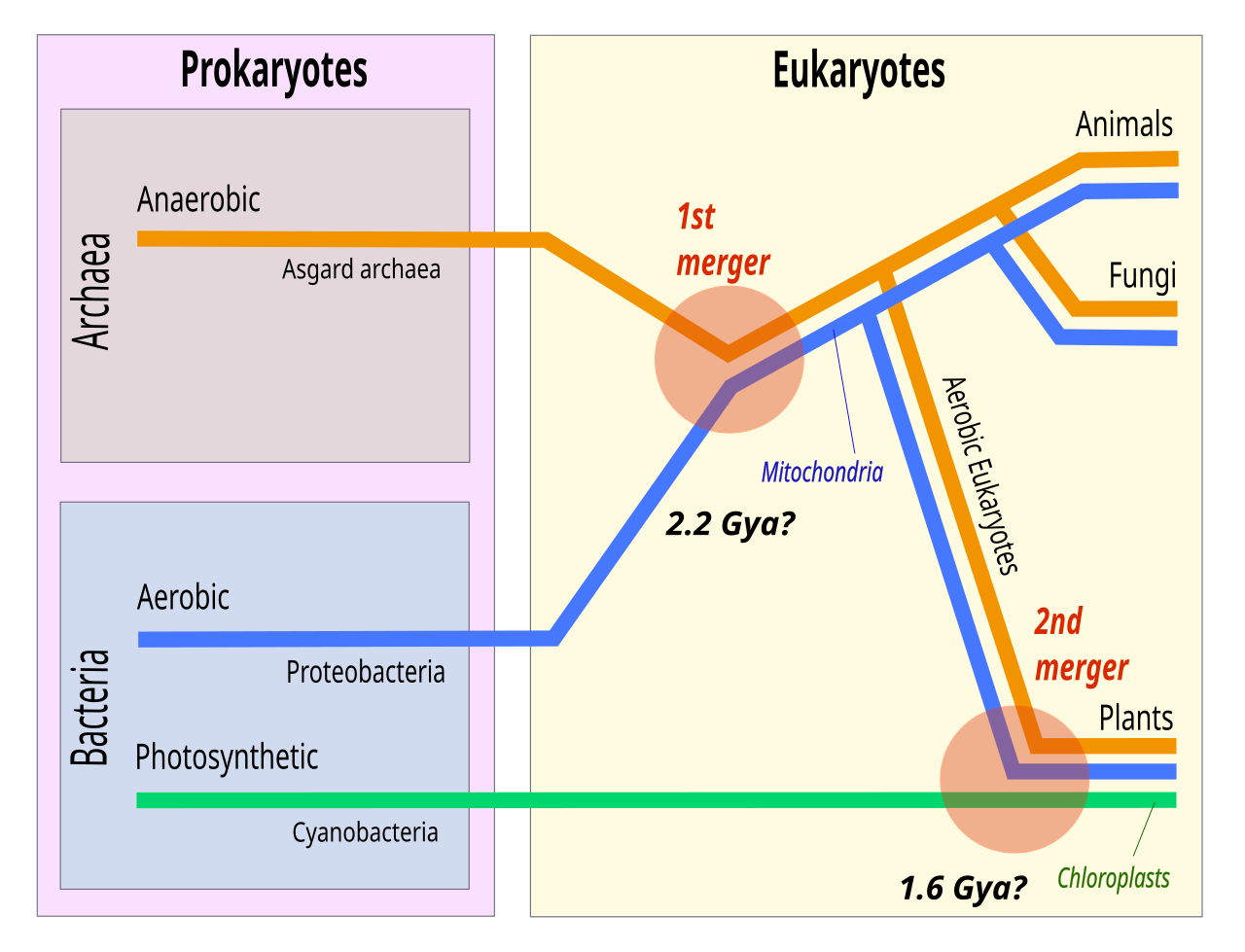
Les facteurs de complication incluent les affirmations selon lesquelles la relation entre les eucaryotes et le phylum archéen Thermoproteota est plus étroite que la relation entre les « Euryarchaeota » et le phylum Thermoproteota et la présence de gènes de type archéens chez certaines bactéries, telles que Thermotoga maritima , issus d' un transfert horizontal de gènes . L'hypothèse standard stipule que l'ancêtre des eucaryotes a divergé tôt des Archaea, et que les eucaryotes sont apparus par symbiogenèse , la fusion d'un archéen et d'une eubactérie, qui a formé les mitochondries ; cette hypothèse explique les similitudes génétiques entre les groupes. L' hypothèse des éocytes postule au contraire que les Eucaryotes ont émergé relativement tard des Archaea.
Une lignée d'archées découverte en 2015, Lokiarchaeum (du nouveau phylum proposé « Lokiarchaeota »), nommée d'après une source hydrothermale appelée Château de Loki dans l'océan Arctique, s'est avérée être la plus proche des eucaryotes connus à cette époque. On l'a qualifiée d'organisme de transition entre les procaryotes et les eucaryotes.
Plusieurs phylums frères de « Lokiarchaeota » ont depuis été découverts (« Thorarchaeota », « Odinarchaeota », « Heimdallarchaeota »), tous constituant ensemble un supergroupe nouvellement proposé Asgard .
Les archées individuelles ont un diamètre compris entre 0,1 micromètre (μm) et plus de 15 μm et se présentent sous diverses formes, généralement sous forme de sphères, de tiges, de spirales ou de plaques. D'autres morphologies chez les Thermoproteota comprennent des cellules lobées de forme irrégulière chez Sulfolobus , des filaments en forme d'aiguilles de moins d'un demi-micromètre de diamètre chez Thermofilum et des tiges presque parfaitement rectangulaires chez Thermoproteus et Pyrobaculum . Les archées du genre Haloquadratum telles que Haloquadratum walsbyi sont des spécimens plats et carrés qui vivent dans des piscines hypersalines. Ces formes inhabituelles sont probablement maintenues à la fois par leurs parois cellulaires et par un cytosquelette procaryote . Des protéines apparentées aux composants du cytosquelette d'autres organismes existent chez les archées, et des filaments se forment à l'intérieur de leurs cellules, mais contrairement à d'autres organismes, ces structures cellulaires sont mal comprises. Chez Thermoplasma et Ferroplasma, l'absence de paroi cellulaire signifie que les cellules ont des formes irrégulières et peuvent ressembler à des amibes .
Certaines espèces forment des agrégats ou des filaments de cellules jusqu'à 200 μm de long. Ces organismes peuvent être importants dans les biofilms . Notamment, les agrégats de cellules de Thermococcus coalescens fusionnent ensemble en culture, formant des cellules géantes uniques. Les archées du genre Pyrodictium produisent une colonie multicellulaire élaborée impliquant des réseaux de tubes creux longs et minces appelés canules qui dépassent de la surface des cellules et les relient en une agglomération dense en forme de buisson. La fonction de ces canules n'est pas établie, mais elles peuvent permettre la communication ou l'échange de nutriments avec les voisins. Il existe des colonies multi-espèces, comme la communauté du « collier de perles » qui a été découverte en 2001 dans un marais allemand. Les colonies rondes et blanchâtres d'une nouvelle espèce d'Euryarchaeota sont espacées le long de minces filaments qui peuvent atteindre 15 centimètres (5,9 pouces) de long ; ces filaments sont constitués d'une espèce bactérienne particulière.
Structure, développement de la composition et fonctionnement
La plupart des archées (mais pas Thermoplasma et Ferroplasma ) possèdent une paroi cellulaire. Chez la plupart des archées, la paroi est assemblée à partir de protéines de la couche de surface, qui forment une couche S. Une couche S est un ensemble rigide de molécules protéiques qui recouvrent l'extérieur de la cellule (comme une cotte de mailles ). [ Cette couche fournit une protection chimique et physique et peut empêcher les macromolécules d'entrer en contact avec la membrane cellulaire. Contrairement aux bactéries, les archées n'ont pas de peptidoglycane dans leurs parois cellulaires. Les méthanobactériennes ont des parois cellulaires contenant du pseudopeptidoglycane , qui ressemble au peptidoglycane eubactérien par sa morphologie, sa fonction et sa structure physique, mais le pseudopeptidoglycane est distinct par sa structure chimique ; il manque d'acides aminés D et d'acide N-acétylmuramique , ce dernier étant remplacé par de l'acide N-acétyltalosaminuronique .
Les flagelles archaïques sont connus sous le nom d' archaella , qui fonctionnent comme les flagelles bactériens - leurs longues tiges sont entraînées par des moteurs rotatifs à la base. Ces moteurs sont alimentés par un gradient de protons à travers la membrane, mais les archaella sont sensiblement différentes en composition et en développement. Les deux types de flagelles ont évolué à partir d'ancêtres différents. Le flagelle bactérien partage un ancêtre commun avec le système de sécrétion de type III , tandis que les flagelles archaïques semblent avoir évolué à partir des pili bactériens de type IV . Contrairement au flagelle bactérien, qui est creux et assemblé par des sous-unités remontant le pore central jusqu'à la pointe du flagelle, les flagelles archaïques sont synthétisés en ajoutant des sous-unités à la base.
Membranes
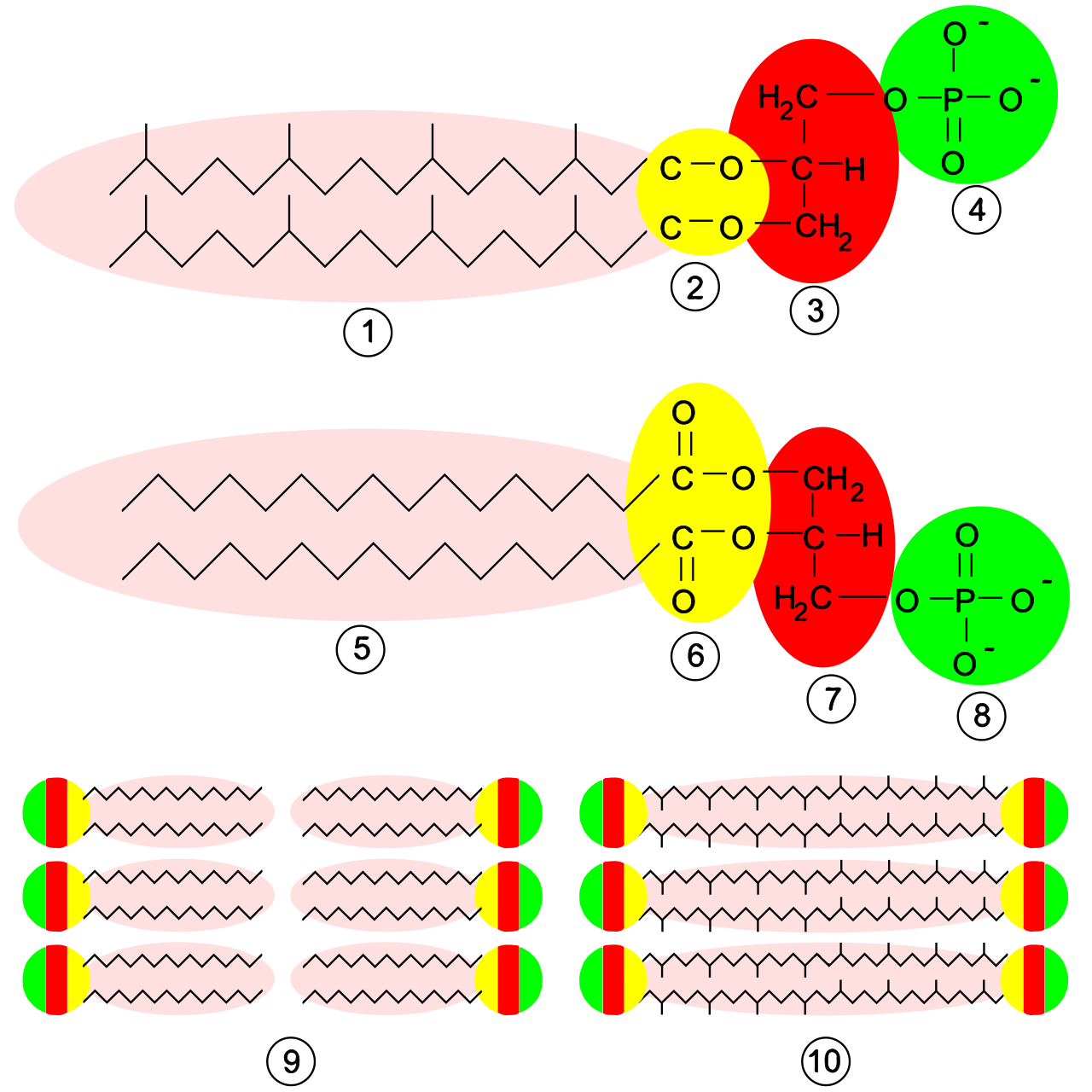
Structures membranaires. En haut , un phospholipide archéen : 1 , chaînes isoprènes ; 2 , liaisons éthers ; 3 , groupement L-glycérol ; 4 , groupement phosphate. Au milieu , un phospholipide bactérien ou eucaryote : 5 , chaînes d'acides gras ; 6 , liaisons esters ; 7 , groupement D-glycérol ; 8 , groupement phosphate. En bas : 9 , bicouche lipidique de bactéries et d'eucaryotes ; 10 , monocouche lipidique de certaines archées.
Les membranes des archées sont constituées de molécules qui sont nettement différentes de celles de toutes les autres formes de vie, ce qui montre que les archées ne sont apparentées que de loin aux bactéries et aux eucaryotes. Chez tous les organismes, les membranes cellulaires sont constituées de molécules appelées phospholipides . Ces molécules possèdent à la fois une partie polaire qui se dissout dans l'eau (la « tête » phosphate ) et une partie non polaire « grasse » qui ne le fait pas (la queue lipidique). Ces parties dissemblables sont reliées par un groupement glycérol . Dans l'eau, les phospholipides se regroupent, les têtes faisant face à l'eau et les queues lui faisant face. La structure principale des membranes cellulaires est une double couche de ces phospholipides, appelée bicouche lipidique .
Les phospholipides des archées sont inhabituels à quatre égards :
Leurs membranes sont composées de lipides glycérol-éther , tandis que les bactéries et les eucaryotes ont des membranes composées principalement de lipides glycérol- ester . La différence réside dans le type de liaison qui relie les lipides à la fraction glycérol ; les deux types sont représentés en jaune sur la figure de droite. Dans les lipides esters, il s'agit d'une liaison ester , tandis que dans les lipides éthers, il s'agit d'une liaison éther .
La stéréochimie de la fraction glycérol des archées est l'image miroir de celle trouvée dans d'autres organismes. La fraction glycérol peut se présenter sous deux formes qui sont des images miroir l'une de l'autre, appelées énantiomères . Tout comme une main droite ne rentre pas facilement dans un gant de gaucher, les énantiomères d'un type ne peuvent généralement pas être utilisés ou fabriqués par des enzymes adaptées à l'autre. Les phospholipides des archées sont construits sur un squelette de sn -glycérol-1-phosphate, qui est un énantiomère de sn -glycérol-3-phosphate, le squelette phospholipidique trouvé dans les bactéries et les eucaryotes. Cela suggère que les archées utilisent des enzymes entièrement différentes pour synthétiser les phospholipides par rapport aux bactéries et aux eucaryotes. De telles enzymes se sont développées très tôt dans l'histoire de la vie, indiquant une séparation précoce des deux autres domaines.
Les queues lipidiques des archées diffèrent de celles des autres organismes en ce qu'elles sont basées sur de longues chaînes isoprénoïdes avec de multiples ramifications latérales, parfois avec des cycles cyclopropane ou cyclohexane . En revanche, les acides gras des membranes d'autres organismes ont des chaînes droites sans ramifications latérales ni cycles. Bien que les isoprénoïdes jouent un rôle important dans la biochimie de nombreux organismes, seules les archées les utilisent pour fabriquer des phospholipides. Ces chaînes ramifiées peuvent aider à empêcher les membranes des archées de fuir à haute température.
Chez certaines archées, la bicouche lipidique est remplacée par une monocouche. En effet, les archées fusionnent les queues de deux molécules de phospholipides en une seule molécule à deux têtes polaires (un bolaamphiphile ) ; cette fusion peut rendre leurs membranes plus rigides et mieux à même de résister aux environnements difficiles. Par exemple, les lipides de Ferroplasma sont de ce type, ce qui est censé favoriser la survie de cet organisme dans son habitat très acide.
D’autres groupes d’archées utilisent la lumière du soleil comme source d’énergie (elles sont phototrophes ), mais la photosynthèse génératrice d’oxygène ne se produit chez aucun de ces organismes. voies métaboliques de base sont communes à toutes les formes de vie ; par exemple, les archées utilisent une forme modifiée de glycolyse (la voie Entner-Doudoroff ) et un cycle complet ou partiel de l’acide citrique . Ces similitudes avec d’autres organismes reflètent probablement à la fois des origines précoces dans l’histoire de la vie et leur haut niveau d’efficacité.
Certains Euryarchaeota sont des méthanogènes (archées qui produisent du méthane à la suite du métabolisme) vivant dans des environnements anaérobies , tels que les marais. Cette forme de métabolisme a évolué tôt, et il est même possible que le premier organisme vivant libre ait été un méthanogène. Une réaction courante implique l'utilisation du dioxyde de carbone comme accepteur d'électrons pour oxyder l'hydrogène . La méthanogénèse implique une gamme de coenzymes qui sont propres à ces archées, telles que la coenzyme M et le méthanofurane . D'autres composés organiques tels que les alcools , l'acide acétique ou l'acide formique sont utilisés comme accepteurs d'électrons alternatifs par les méthanogènes. Ces réactions sont courantes chez les archées vivant dans l'intestin . L'acide acétique est également décomposé directement en méthane et en dioxyde de carbone, par les archées acétotrophes . Ces acétotrophes sont des archées de l'ordre des Methanosarcinales et constituent une part importante des communautés de micro-organismes qui produisent du biogaz .
D'autres archées utilisent le CO 2Les archées se fixent dans l' atmosphère comme source de carbone, dans un processus appelé fixation du carbone (elles sont autotrophes ). Ce processus implique soit une forme hautement modifiée du cycle de Calvin , soit une autre voie métabolique appelée cycle 3-hydroxypropionate/4-hydroxybutyrate. Les Thermoproteota utilisent également le cycle de Krebs inverse tandis que les « Euryarchaeota » utilisent également la voie réductrice de l'acétyl-CoA . La fixation du carbone est alimentée par des sources d'énergie inorganiques. Aucune archée connue ne pratique la photosynthèse ( Halobacterium est le seul archéon phototrophe connu, mais il utilise un processus alternatif à la photosynthèse). Les sources d'énergie des archées sont extrêmement diverses et vont de l'oxydation de l'ammoniac par les Nitrosopumilales à l'oxydation du sulfure d'hydrogène ou du soufre élémentaire par des espèces de Sulfolobus , en utilisant soit de l'oxygène, soit des ions métalliques comme accepteurs d'électrons.
Les archées phototrophes utilisent la lumière pour produire de l'énergie chimique sous forme d'ATP. Chez les Halobactéries , des pompes à ions activées par la lumière comme la bactériorhodopsine et l'halorhodopsine génèrent des gradients ioniques en pompant les ions hors et dans la cellule à travers la membrane plasmique . L'énergie stockée dans ces gradients électrochimiques est ensuite convertie en ATP par l'ATP synthase . Ce processus est une forme de photophosphorylation . La capacité de ces pompes à ions activées par la lumière à déplacer les ions à travers les membranes dépend des changements induits par la lumière dans la structure d'un cofacteur de rétinol enfoui au centre de la protéine.
Génétique
Les archées ont généralement un seul chromosome circulaire , mais il a été démontré que de nombreuses euryarchées portent plusieurs copies de ce chromosome. Le plus grand génome archéen connu en 2002 était de 5 751 492 paires de bases chez Methanosarcina acetivorans . Le minuscule génome de 490 885 paires de bases de Nanoarchaeum equitans est un dixième de cette taille et le plus petit génome archéen connu ; on estime qu'il ne contient que 537 gènes codant pour des protéines. Des fragments d'ADN indépendants plus petits, appelés plasmides , sont également présents chez les archées. Les plasmides peuvent être transférés entre les cellules par contact physique, dans un processus qui peut être similaire à la conjugaison bactérienne .
Les archées sont génétiquement distinctes des bactéries et des eucaryotes, avec jusqu'à 15 % des protéines codées par un génome archéen étant uniques au domaine, bien que la plupart de ces gènes uniques n'aient pas de fonction connue. Parmi les protéines uniques restantes qui ont une fonction identifiée, la plupart appartiennent aux Euryarchaeota et sont impliquées dans la méthanogénèse. Les protéines que partagent les archées, les bactéries et les eucaryotes forment un noyau commun de fonction cellulaire, principalement lié à la transcription , à la traduction et au métabolisme des nucléotides . D'autres caractéristiques archéennes sont l'organisation de gènes de fonction apparentée - tels que les enzymes qui catalysent les étapes de la même voie métabolique en nouveaux opérones , et de grandes différences dans les gènes d'ARNt et leurs aminoacyl ARNt synthétases .
Français La transcription chez les archées ressemble plus étroitement à la transcription eucaryote qu'à la transcription bactérienne, l' ARN polymérase archéenne étant très proche de son équivalent eucaryote, tandis que la traduction archéenne montre des signes d'équivalents bactériens et eucaryotes. Bien que les archées n'aient qu'un seul type d'ARN polymérase, sa structure et sa fonction dans la transcription semblent être proches de celles de l' ARN polymérase II eucaryote , avec des assemblages de protéines similaires (les facteurs de transcription généraux ) dirigeant la liaison de l'ARN polymérase au promoteur d'un gène , mais d'autres facteurs de transcription archaïques sont plus proches de ceux trouvés dans les bactéries. La modification post-transcriptionnelle est plus simple que chez les eucaryotes, car la plupart des gènes archaïques n'ont pas d'introns , bien qu'il y ait de nombreux introns dans leurs gènes d'ARN de transfert et d'ARN ribosomique , et des introns peuvent apparaître dans quelques gènes codant pour des protéines.
Transfert de gènes et échange génétique
Haloferax volcanii , un archéon halophile extrême, forme des ponts cytoplasmiques entre les cellules qui semblent être utilisés pour le transfert d'ADN d'une cellule à une autre dans les deux sens.
Français Lorsque les archées hyperthermophiles Sulfolobus solfataricus et Sulfolobus acidocaldarius sont exposées à une irradiation UV endommageant l'ADN ou aux agents bléomycine ou mitomycine C , une agrégation cellulaire spécifique à l'espèce est induite. L'agrégation chez S. solfataricus n'a pas pu être induite par d'autres facteurs de stress physiques, tels que le pH ou le changement de température, ce qui suggère que l'agrégation est induite spécifiquement par des dommages à l'ADN . Ajon et al. ont montré que l'agrégation cellulaire induite par les UV médie l'échange de marqueurs chromosomiques avec une fréquence élevée chez S. acidocaldarius . Les taux de recombinaison ont dépassé ceux des cultures non induites jusqu'à trois ordres de grandeur. Frols et al. et Ajon et al. ont émis l'hypothèse que l'agrégation cellulaire améliore le transfert d'ADN spécifique à l'espèce entre les cellules Sulfolobus afin de permettre une réparation accrue de l'ADN endommagé au moyen de la recombinaison homologue . Cette réponse peut être une forme primitive d'interaction sexuelle similaire aux systèmes de transformation bactérienne les mieux étudiés qui sont également associés au transfert d'ADN spécifique à l'espèce entre les cellules conduisant à la réparation par recombinaison homologue des dommages à l'ADN.
Virus archaïques
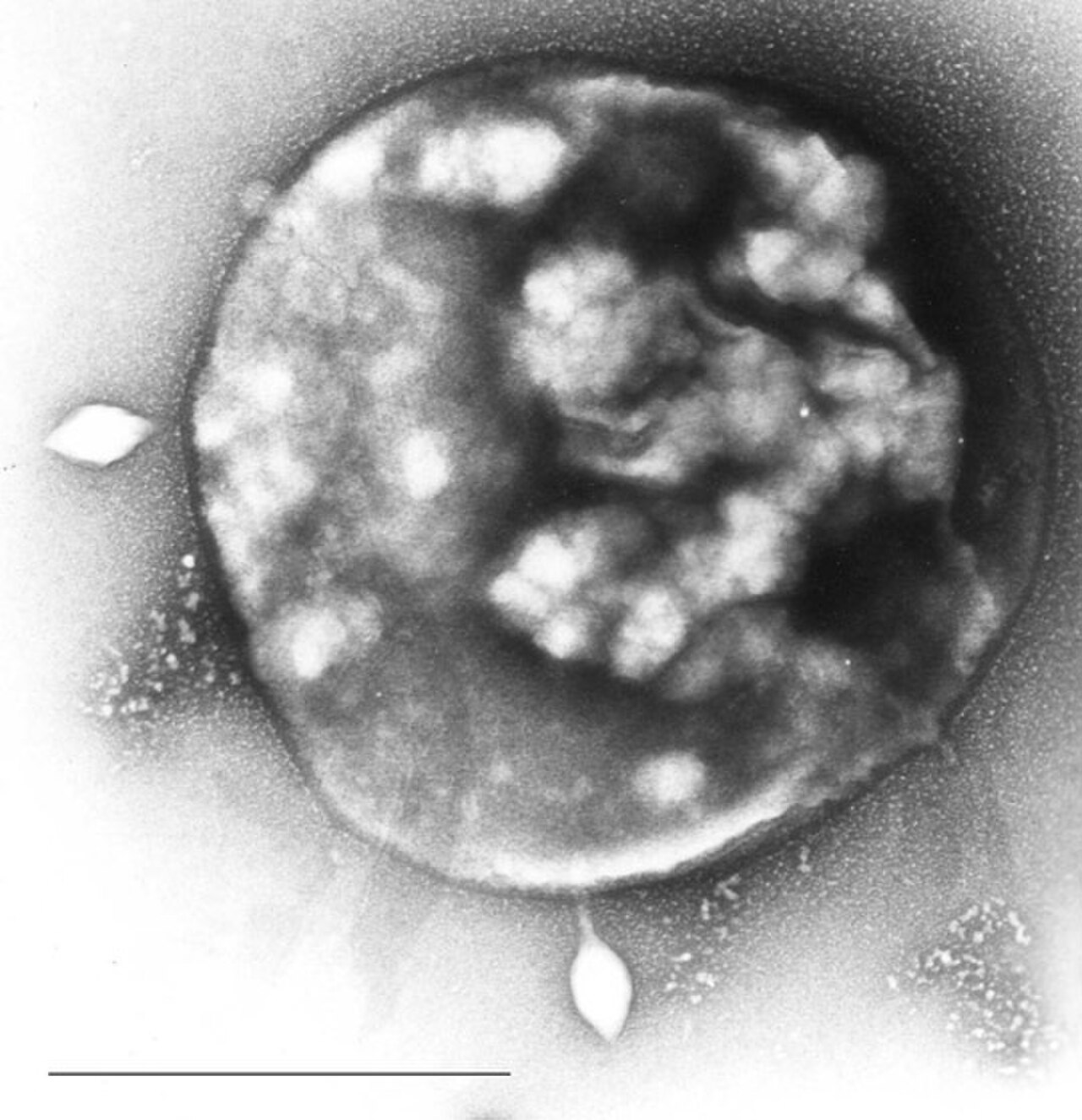
Les archées sont la cible d'un certain nombre de virus dans une virosphère diversifiée distincte des virus bactériens et eucaryotes. Elles ont été organisées en 15 à 18 familles basées sur l'ADN jusqu'à présent, mais de nombreuses espèces restent non isolées et attendent d'être classées. Ces familles peuvent être divisées de manière informelle en deux groupes : spécifiques aux archées et cosmopolites. Les virus spécifiques aux archées ne ciblent que les espèces archéennes et comprennent actuellement 12 familles. De nombreuses structures virales uniques, non identifiées auparavant, ont été observées dans ce groupe, notamment : des virus en forme de bouteille, en forme de fuseau, en forme de bobine et en forme de gouttelettes. Bien que les cycles de reproduction et les mécanismes génomiques des espèces spécifiques aux archées puissent être similaires à ceux d'autres virus, ils présentent des caractéristiques uniques qui ont été spécifiquement développées en raison de la morphologie des cellules hôtes qu'ils infectent. Leurs mécanismes de libération du virus diffèrent de ceux des autres phages. Les bactériophages suivent généralement soit des voies lytiques , soit des voies lysogènes , soit (rarement) un mélange des deux. La plupart des souches virales spécifiques aux archées entretiennent une relation stable, quelque peu lysogène, avec leurs hôtes, se manifestant par une infection chronique. Cela implique la production et la libération progressives et continues de virions sans tuer la cellule hôte. Prangishyili (2013) a noté qu'il a été émis l'hypothèse que les phages archaïques à queue provenaient de bactériophages capables d'infecter les espèces haloarchéennes . Si l'hypothèse est correcte, on peut conclure que les autres virus à ADN double brin qui constituent le reste du groupe spécifique aux archées constituent leur propre groupe unique dans la communauté virale mondiale. Krupovic et al. (2018) déclarent que les niveaux élevés de transfert horizontal de gènes , les taux de mutation rapides dans les génomes viraux et l'absence de séquences génétiques universelles ont conduit les chercheurs à percevoir la voie évolutive des virus archaïques comme un réseau. L'absence de similitudes entre les marqueurs phylogénétiques de ce réseau et de la virosphère globale, ainsi que les liens externes avec des éléments non viraux, peuvent suggérer que certaines espèces de virus spécifiques aux archées ont évolué à partir d'éléments génétiques mobiles non viraux (MGE).
Ces virus ont été étudiés en détail chez les thermophiles, en particulier les ordres Sulfolobales et Thermoproteales . Deux groupes de virus à ADN simple brin qui infectent les archées ont été récemment isolés. L'un de ces groupes est illustré par le virus pléomorphe Halorubrum 1 ( Pleolipoviridae ) qui infecte les archées halophiles, et l'autre par le virus en forme de bobine Aeropyrum ( Spiraviridae ) qui infecte un hôte hyperthermophile (croissance optimale à 90–95 °C). Il convient de noter que ce dernier virus possède le plus grand génome à ADN simple brin actuellement signalé. Les défenses contre ces virus peuvent impliquer une interférence ARN à partir de séquences d'ADN répétitives qui sont liées aux gènes des virus.
Reproduction
Les archées se reproduisent de manière asexuée par fission binaire ou multiple , fragmentation ou bourgeonnement ; la mitose et la méiose ne se produisent pas, donc si une espèce d'archées existe sous plus d'une forme, toutes ont le même matériel génétique. La division cellulaire est contrôlée dans un cycle cellulaire ; après que le chromosome de la cellule est répliqué et que les deux chromosomes filles se séparent, la cellule se divise. Dans le genre Sulfolobus , le cycle a des caractéristiques similaires à celles des systèmes bactériens et eucaryotes. Les chromosomes se répliquent à partir de plusieurs points de départ ( origines de réplication ) en utilisant des ADN polymérases qui ressemblent aux enzymes eucaryotes équivalentes.
Chez les Euryarchaeota, la protéine de division cellulaire FtsZ , qui forme un anneau de contraction autour de la cellule, et les composants du septum construit au centre de la cellule, sont similaires à leurs équivalents bactériens. Chez les cren- et les thaumarchaea, le mécanisme de division cellulaire Cdv remplit un rôle similaire. Ce mécanisme est lié au mécanisme eucaryote ESCRT-III qui, bien que mieux connu pour son rôle dans le tri cellulaire, a également été vu comme jouant un rôle dans la séparation entre les cellules divisées, suggérant un rôle ancestral dans la division cellulaire.
Les bactéries et les eucaryotes, mais pas les archées, produisent des spores . Certaines espèces de Haloarchaea subissent une commutation phénotypique et se développent sous forme de plusieurs types de cellules différents, y compris des structures à parois épaisses qui résistent au choc osmotique et permettent aux archées de survivre dans l'eau à de faibles concentrations de sel, mais ce ne sont pas des structures reproductrices et peuvent plutôt les aider à atteindre de nouveaux habitats.
Comportement
Communication
À l'origine, on pensait que la détection de quorum n'existait pas chez les archées, mais des études récentes ont montré que certaines espèces étaient capables d'effectuer des échanges par détection de quorum. D'autres études ont montré des interactions syntrophiques entre les archées et les bactéries pendant la croissance du biofilm. Bien que la recherche sur la détection de quorum chez les archées soit limitée, certaines études ont découvert des protéines LuxR chez les espèces archéennes, présentant des similitudes avec la LuxR des bactéries, et permettant finalement la détection de petites molécules utilisées dans la communication à haute densité. Comme pour les bactéries, les solos LuxR des archées se lient aux AHL (lactones) et aux ligans non-AHL, ce qui joue un rôle important dans la réalisation de communications intra-espèces, inter-espèces et inter-règnes par détection de quorum.
Les archées extrémophiles appartiennent à quatre groupes physiologiques principaux : les halophiles , les thermophiles , les alcalinophiles et les acidophiles . Ces groupes ne sont pas exhaustifs ni spécifiques à un embranchement, et ne s'excluent pas mutuellement, car certaines archées appartiennent à plusieurs groupes. Néanmoins, ils constituent un point de départ utile pour la classification.
Les halophiles, dont le genre Halobacterium , vivent dans des environnements extrêmement salins tels que les lacs salés et sont plus nombreux que leurs homologues bactériens à des salinités supérieures à 20–25 %. Les thermophiles se développent mieux à des températures supérieures à 45 °C (113 °F), dans des endroits tels que les sources chaudes ; les archées hyperthermophiles se développent de manière optimale à des températures supérieures à 80 °C (176 °F). L'archée Methanopyrus kandleri souche 116 peut même se reproduire à 122 °C (252 °F), la température la plus élevée enregistrée de tous les organismes.
D'autres archées existent dans des conditions très acides ou alcalines. Par exemple, l'un des acidophiles archéens les plus extrêmes est Picrophilus torridus , qui pousse à un pH de 0, ce qui équivaut à prospérer dans de l'acide sulfurique 1,2 molaire .
Cette résistance aux environnements extrêmes a fait des archées l'objet de spéculations sur les propriétés possibles de la vie extraterrestre . Certains habitats extrémophiles ne sont pas différents de ceux de Mars , ce qui conduit à suggérer que des microbes viables pourraient être transférés entre les planètes dans les météorites .
Récemment, plusieurs études ont montré que les archées n'existent pas seulement dans les environnements mésophiles et thermophiles, mais sont également présentes, parfois en grand nombre, à basse température. Par exemple, les archées sont courantes dans les environnements océaniques froids tels que les mers polaires. Plus significatif encore est le grand nombre d'archées trouvées dans tous les océans du monde dans des habitats non extrêmes au sein de la communauté planctonique (en tant que partie du picoplancton ). Bien que ces archées puissent être présentes en nombre extrêmement élevé (jusqu'à 40 % de la biomasse microbienne), presque aucune de ces espèces n'a été isolée et étudiée en culture pure . Par conséquent, notre compréhension du rôle des archées dans l'écologie océanique est rudimentaire, de sorte que leur pleine influence sur les cycles biogéochimiques mondiaux reste largement inexplorée. Certains Thermoproteota marins sont capables de nitrification , ce qui suggère que ces organismes peuvent affecter le cycle de l'azote océanique , bien que ces Thermoproteota océaniques puissent également utiliser d'autres sources d'énergie.
On trouve également un grand nombre d'archées dans les sédiments qui recouvrent le fond marin , ces organismes constituant la majorité des cellules vivantes à des profondeurs supérieures à 1 mètre sous le fond de l'océan. Il a été démontré que dans tous les sédiments de surface océaniques (de 1 000 à 10 000 m de profondeur d'eau), l'impact de l'infection virale est plus élevé sur les archées que sur les bactéries et la lyse des archées induite par le virus représente jusqu'à un tiers de la biomasse microbienne totale tuée, ce qui entraîne la libération d'environ 0,3 à 0,5 gigatonne de carbone par an à l'échelle mondiale.
Rôle dans le cycle chimique
Les archées recyclent des éléments tels que le carbone , l'azote et le soufre à travers leurs différents habitats. Les archées effectuent de nombreuses étapes du cycle de l'azote . Cela comprend à la fois des réactions qui éliminent l'azote des écosystèmes (comme la respiration à base de nitrate et la dénitrification ) ainsi que des processus qui introduisent de l'azote (comme l'assimilation des nitrates et la fixation de l'azote ).
Des chercheurs ont récemment découvert l'implication des archées dans les réactions d'oxydation de l'ammoniac . Ces réactions sont particulièrement importantes dans les océans. Les archées semblent également cruciales pour l'oxydation de l'ammoniac dans les sols. Elles produisent du nitrite , que d'autres microbes oxydent ensuite en nitrate . Les plantes et d'autres organismes consomment ce dernier.
Dans le cycle du soufre , les archées qui se développent en oxydant les composés soufrés libèrent cet élément des roches, le rendant ainsi disponible pour d'autres organismes, mais les archées qui font cela, comme Sulfolobus , produisent de l'acide sulfurique comme déchet, et la croissance de ces organismes dans les mines abandonnées peut contribuer au drainage minier acide et à d'autres dommages environnementaux.
Dans le cycle du carbone , les archées méthanogènes éliminent l'hydrogène et jouent un rôle important dans la décomposition de la matière organique par les populations de micro-organismes qui agissent comme décomposeurs dans les écosystèmes anaérobies, tels que les sédiments, les marais et les stations d'épuration .
Interactions avec d'autres organismes
Les archées méthanogènes forment une symbiose avec les termites , vivant dans leur intestin et aidant à digérer la cellulose.
Les interactions bien caractérisées entre les archées et d'autres organismes sont soit mutuelles , soit commensales . Il n'existe pas d'exemples clairs d' agents pathogènes ou de parasites archéens connus , mais certaines espèces de méthanogènes ont été suggérées comme étant impliquées dans des infections de la bouche , et Nanoarchaeum equitans pourrait être un parasite d'une autre espèce d'archées, puisqu'il ne survit et ne se reproduit que dans les cellules du Crenarchaeon Ignicoccus hospitalis , et ne semble offrir aucun avantage à son hôte .
Mutualisme
Le mutualisme est une interaction entre des individus de différentes espèces qui entraîne des effets positifs (bénéfiques) sur la reproduction par habitant et/ou la survie des populations en interaction. Un exemple bien connu de mutualisme est l'interaction entre les protozoaires et les archées méthanogènes dans le tube digestif des animaux qui digèrent la cellulose , comme les ruminants et les termites . Dans ces environnements anaérobies, les protozoaires décomposent la cellulose végétale pour obtenir de l'énergie. Ce processus libère de l'hydrogène comme déchet, mais des niveaux élevés d'hydrogène réduisent la production d'énergie. Lorsque les méthanogènes convertissent l'hydrogène en méthane, les protozoaires bénéficient de plus d'énergie.
Chez les protozoaires anaérobies, tels que Plagiopyla frontata , Trimyema , Heterometopus et Metopus contortus , les archées résident à l'intérieur des protozoaires et consomment l'hydrogène produit dans leurs hydrogénosomes . Les archées s'associent également à des organismes plus grands. Par exemple, l'archée marine Cenarchaeum symbiosum est un endosymbiote de l' éponge Axinella mexicana .
Commensalisme
Certaines archées sont commensales, c'est-à-dire qu'elles bénéficient d'une association sans aider ni nuire à l'autre organisme. Par exemple, le méthanogène Methanobrevibacter smithii est de loin l'archée le plus commun dans la flore humaine , représentant environ un procaryote sur dix dans l'intestin humain. Chez les termites et chez l'homme, ces méthanogènes peuvent en fait être mutualistes, interagissant avec d'autres microbes dans l'intestin pour faciliter la digestion. Les communautés archéennes s'associent à toute une gamme d'autres organismes, comme à la surface des coraux , et dans la région du sol qui entoure les racines des plantes (la rhizosphère ).
Parasitisme
Bien que les archées n'aient pas la réputation historique d'être des agents pathogènes, on les trouve souvent avec des génomes similaires à ceux d'agents pathogènes plus courants comme E. coli, montrant des liens métaboliques et une histoire évolutive avec les agents pathogènes actuels. Les archées ont été détectées de manière incohérente dans les études cliniques en raison du manque de catégorisation des archées en espèces plus spécifiques.
Les archées hébergent une nouvelle classe d' antibiotiques potentiellement utiles . Quelques-unes de ces archéocines ont été caractérisées, mais on pense qu'il en existe des centaines d'autres, en particulier dans les genres Haloarchaea et Sulfolobus . Ces composés diffèrent par leur structure des antibiotiques bactériens, de sorte qu'ils pourraient avoir de nouveaux modes d'action. En outre, ils pourraient permettre la création de nouveaux marqueurs sélectionnables à utiliser en biologie moléculaire des archées.
.jpg/1280px-Aerial_image_of_Grand_Prismatic_Spring_(view_from_the_south).jpg)