Les éponges ou éponges de mer sont des invertébrés marins de l' embranchement des métazoaires Porifera ( / p ə ˈ r ɪ f ər ə ˌ p ɔː -/ pər- IF -ər-ə, por- ; signifiant « porteur ...
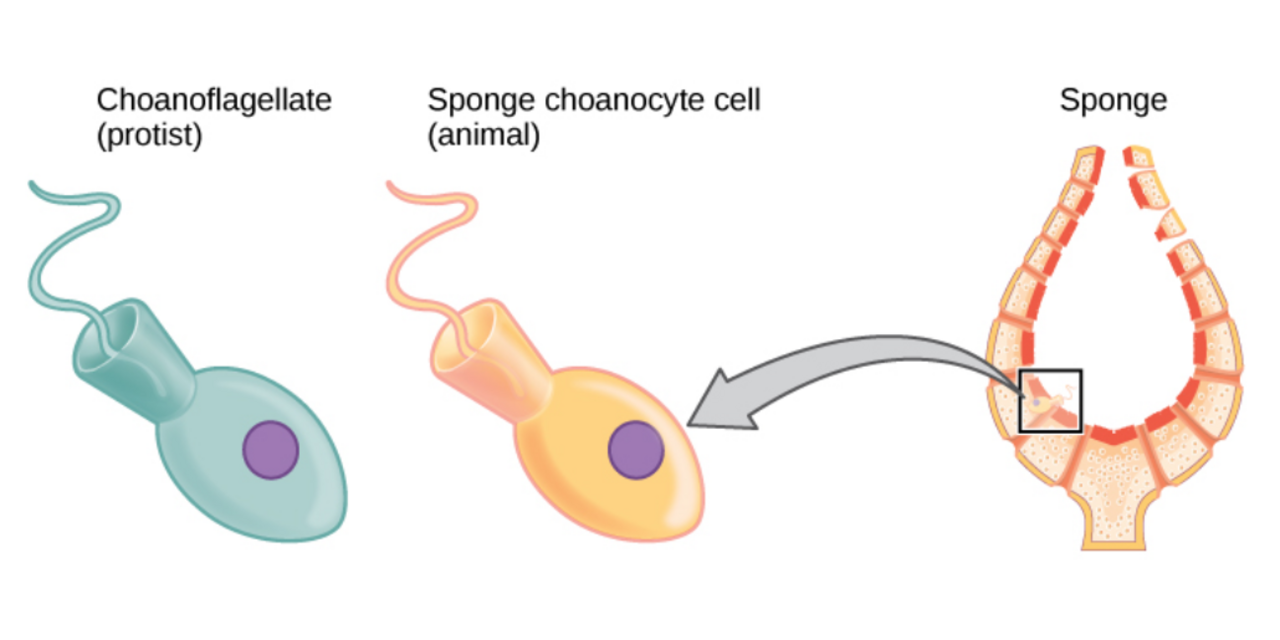
Les éponges sont semblables aux autres animaux en ce qu'elles sont multicellulaires , hétérotrophes , dépourvues de parois cellulaires et produisent des spermatozoïdes . Contrairement aux autres animaux, elles n'ont pas de véritables tissus et organes ]. Certaines d'entre elles sont radialement symétriques, mais la plupart sont asymétriques. Les formes de leur corps sont adaptées pour une efficacité maximale de l'écoulement de l'eau à travers la cavité centrale, où l'eau dépose les nutriments puis sort par un trou appelé osculum . Les choanoflagellés unicellulaires ressemblent aux cellules choanocytes des éponges qui sont utilisées pour piloter leurs systèmes d'écoulement de l'eau et capturer la plupart de leur nourriture. Ceci, ainsi que des études phylogénétiques des molécules ribosomiques, ont été utilisées comme preuve morphologique pour suggérer que les éponges sont le groupe frère du reste des animaux . Une grande majorité sont des espèces marines (eau salée), dont l'habitat s'étend des zones de marée à des profondeurs dépassant 8 800 m (5,5 mi), bien qu'il existe des espèces d'eau douce. Toutes les éponges adultes sont sessiles , ce qui signifie qu'elles se fixent à une surface sous-marine et restent en place (c'est-à-dire qu'elles ne se déplacent pas). Au stade larvaire , elles sont mobiles .
De nombreuses éponges ont des squelettes internes de spicules (fragments de carbonate de calcium ou de dioxyde de silicium ressemblant à des squelettes ) et/ou de spongine (un type modifié de protéine de collagène). Une matrice gélatineuse interne appelée mésohyle fonctionne comme un endosquelette et c'est le seul squelette des éponges molles qui encroûtent des surfaces dures comme les rochers. Le plus souvent, le mésohyle est rigidifié par des spicules minéraux , par des fibres de spongine ou les deux. 90 % de toutes les espèces d'éponges connues qui ont la plus large gamme d'habitats, y compris toutes celles d'eau douce, sont des démosponges qui utilisent la spongine ; de nombreuses espèces ont des spicules de silice , tandis que certaines espèces ont des exosquelettes de carbonate de calcium . Les éponges calcaires ont des spicules de carbonate de calcium et, chez certaines espèces, des exosquelettes de carbonate de calcium, sont limitées aux eaux marines relativement peu profondes où la production de carbonate de calcium est la plus facile. Les éponges de verre fragiles , avec un « échafaudage » de spicules de silice, sont limitées aux régions polaires et aux profondeurs océaniques où les prédateurs sont rares. Des fossiles de tous ces types ont été trouvés dans des roches datées de 580 millions d'années . De plus, les archéocyathides , dont les fossiles sont courants dans les roches datant de 530 à 490 millions d'années , sont désormais considérés comme un type d'éponge.
Bien que la plupart des 5 000 à 10 000 espèces d’éponges connues se nourrissent de bactéries et d’autres aliments microscopiques présents dans l’eau, certaines hébergent des micro-organismes photosynthétiques en tant qu’endosymbiotes , et ces alliances produisent souvent plus de nourriture et d’oxygène qu’elles n’en consomment. Quelques espèces d’éponges qui vivent dans des environnements pauvres en nourriture ont évolué en carnivores qui se nourrissent principalement de petits crustacés .
La plupart des éponges se reproduisent de manière sexuée , mais elles peuvent aussi se reproduire de manière asexuée. Les espèces à reproduction sexuée libèrent des spermatozoïdes dans l'eau pour fertiliser les ovules libérés ou retenus par leur partenaire ou « mère » ; les œufs fécondés se développent en larves qui nagent à la recherche d'endroits où s'installer. Les éponges sont connues pour se régénérer à partir de fragments cassés, bien que cela ne fonctionne que si les fragments contiennent les bons types de cellules. Certaines espèces se reproduisent par bourgeonnement. Lorsque les conditions environnementales deviennent moins hospitalières pour les éponges, par exemple lorsque les températures baissent, de nombreuses espèces d'eau douce et quelques espèces marines produisent des gemmules , des « gousses de survie » de cellules non spécialisées qui restent dormantes jusqu'à ce que les conditions s'améliorent ; elles forment alors des éponges complètement nouvelles ou recolonisent les squelettes de leurs parents.
Les cellules du clade protiste des choanoflagellés ressemblent beaucoup aux cellules des choanocytes d'éponge. Le battement des flagelles des choanocytes attire l'eau à travers l'éponge afin que les nutriments puissent être extraits et les déchets éliminés.
Les quelques espèces de démosponges qui ont un squelette entièrement fibreux et mou, sans éléments durs, ont été utilisées par les humains pendant des milliers d'années à diverses fins, notamment comme rembourrage et comme outils de nettoyage. Dans les années 1950, cependant, ces espèces ont été surexploitées à tel point que l'industrie s'est presque effondrée, et la plupart des matériaux ressemblant à des éponges sont désormais synthétiques. Les éponges et leurs endosymbiotes microscopiques font maintenant l'objet de recherches en tant que sources possibles de médicaments pour traiter un large éventail de maladies. Des dauphins ont été observés en train d'utiliser des éponges comme outils pour chercher de la nourriture .
Caractéristiques distinctives
Les éponges constituent le phylum Porifera et ont été définies comme des métazoaires sessiles (animaux immobiles multicellulaires) qui ont des ouvertures d'admission et de sortie d'eau reliées par des chambres tapissées de choanocytes , des cellules avec des flagelles en forme de fouet. Cependant, quelques éponges carnivores ont perdu ces systèmes d'écoulement d'eau et les choanocytes. Toutes les éponges vivantes connues peuvent remodeler leur corps, car la plupart des types de leurs cellules peuvent se déplacer dans leur corps et quelques-unes peuvent passer d'un type à un autre.
Même si quelques éponges sont capables de produire du mucus – qui agit comme une barrière microbienne chez tous les autres animaux – aucune éponge capable de sécréter une couche de mucus fonctionnelle n’a été répertoriée. Sans une telle couche de mucus, leurs tissus vivants sont recouverts d’une couche de symbiotes microbiens, qui peuvent représenter jusqu’à 40 à 50 % de la masse humide de l’éponge. Cette incapacité à empêcher les microbes de pénétrer dans leur tissu poreux pourrait être l’une des principales raisons pour lesquelles elles n’ont jamais développé une anatomie plus complexe.
Comme les cnidaires (méduses, etc.) et les cténophores (méduses à peigne), et contrairement à tous les autres métazoaires connus, le corps des éponges est constitué d'une masse gélatineuse non vivante ( mésohyle ) coincée entre deux couches principales de cellules. Les cnidaires et les cténophores ont des systèmes nerveux simples, et leurs couches cellulaires sont liées par des connexions internes et en étant montées sur une membrane basale (mince tapis fibreux, également connu sous le nom de « lame basale »). Les éponges n'ont pas de système nerveux similaire à celui des vertébrés, mais peuvent en avoir un qui est assez différent. Leurs couches gélatineuses médianes ont des populations de cellules importantes et variées, et certains types de cellules de leurs couches externes peuvent se déplacer dans la couche médiane et changer de fonction.
Structure de base
Types de cellules
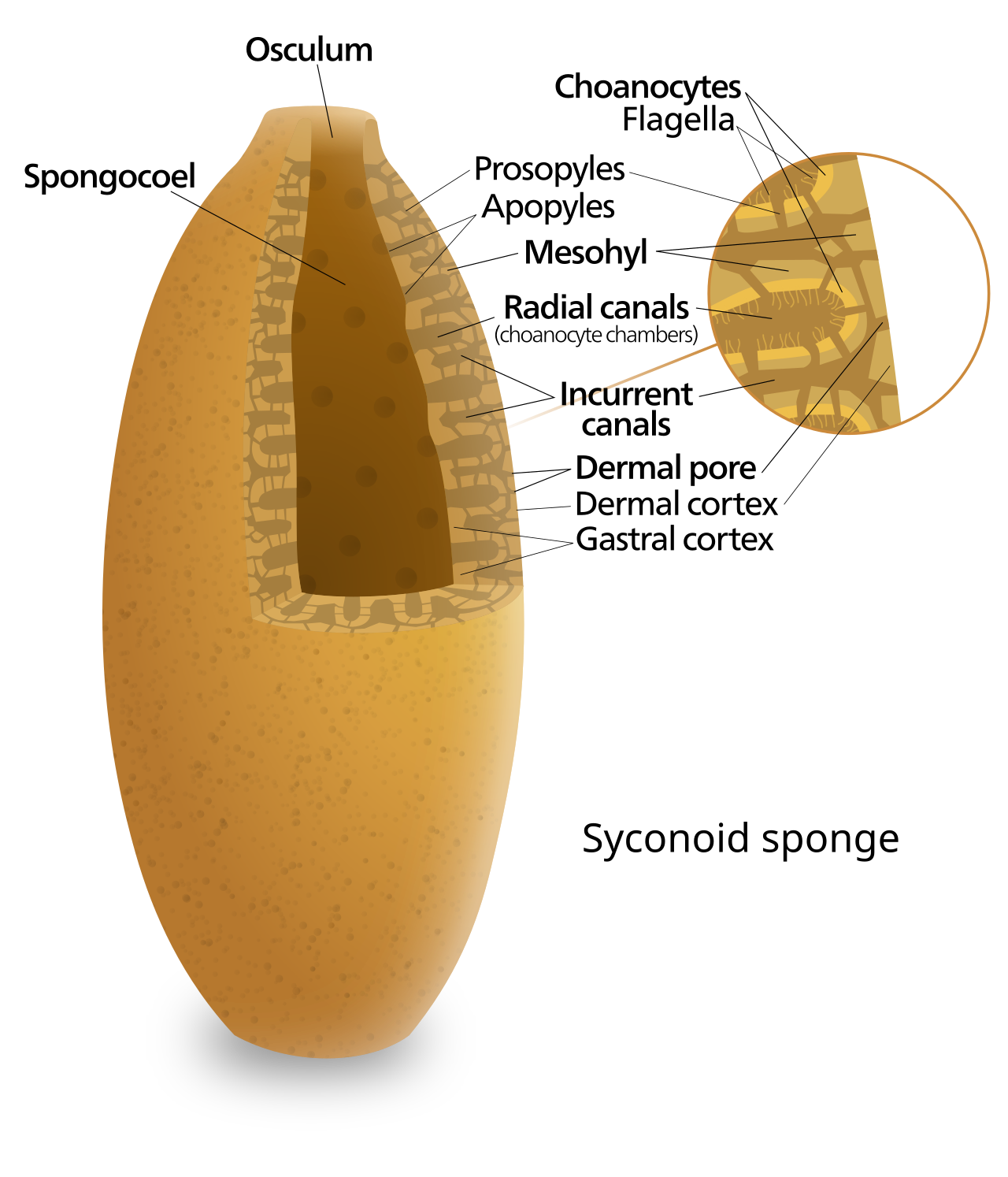
Le corps d'une éponge est creux et maintenu en forme par le mésohyle , une substance gélatineuse composée principalement de collagène et renforcée par un réseau dense de fibres également constituées de collagène. 18 types de cellules distincts ont été identifiés. La surface interne est recouverte de choanocytes , des cellules à colliers cylindriques ou coniques entourant un flagelle par choanocyte. Le mouvement ondulatoire des flagelles en forme de fouet propulse l'eau à travers le corps de l'éponge. Toutes les éponges ont des ostia , des canaux menant à l'intérieur à travers le mésohyle, et dans la plupart des éponges, ceux-ci sont contrôlés par des porocytes en forme de tube qui forment des valves d'entrée fermables. Les pinacocytes , des cellules en forme de plaque, forment une peau externe monocouche sur toutes les autres parties du mésohyle qui ne sont pas recouvertes de choanocytes, et les pinacocytes digèrent également les particules alimentaires trop grosses pour entrer dans les ostia, tandis que ceux à la base de l'animal sont responsables de son ancrage.
D'autres types de cellules vivent et se déplacent dans le mésohyle :
Les lophocytes sont des cellules semblables à des amibes qui se déplacent lentement dans le mésohyle et sécrètent des fibres de collagène.
Les collencytes sont un autre type de cellules productrices de collagène.
Les cellules rhabdifères sécrètent des polysaccharides qui font également partie du mésohyle.
Les sclérocytes sécrètent les spicules minéralisés (« petites épines ») qui forment le squelette de nombreuses éponges et qui, chez certaines espèces, assurent une certaine défense contre les prédateurs.
En plus ou à la place des sclérocytes, les démosponges possèdent des spongocytes qui sécrètent une forme de collagène qui se polymérise en spongine , un matériau fibreux épais qui rigidifie le mésohyle.
Les myocytes (« cellules musculaires ») conduisent des signaux et provoquent la contraction de certaines parties de l’animal.
Les « cellules grises » agissent comme des éponges, l’équivalent d’un système immunitaire .
Les archéocytes (ou amoebocytes ) sont des cellules de type amibien qui sont totipotentes , c'est-à-dire qu'elles sont chacune capables de se transformer en n'importe quel autre type de cellule. Elles jouent également un rôle important dans l'alimentation et dans l'élimination des débris qui bloquent les ostia.
La plupart des éponges fonctionnent un peu comme des cheminées : elles absorbent l'eau par le bas et l'éjectent par l' oscule au sommet. Comme les courants ambiants sont plus rapides au sommet, l'effet de succion qu'elles produisent selon le principe de Bernoulli fait une partie du travail gratuitement. Les éponges peuvent contrôler le flux d'eau en fermant partiellement ou totalement l'oscule et les ostia (les pores d'admission) et en faisant varier le rythme des flagelles, et peuvent même l'arrêter s'il y a beaucoup de sable ou de limon dans l'eau.
Bien que les couches de pinacocytes et de choanocytes ressemblent aux épithéliums d'animaux plus complexes, elles ne sont pas étroitement liées par des connexions intercellulaires ou par une lame basale (une fine feuille fibreuse en dessous). La flexibilité de ces couches et le remodelage du méso-hyle par les lophocytes permettent aux animaux d'ajuster leurs formes tout au long de leur vie pour tirer le meilleur parti des courants d'eau locaux.
La structure corporelle la plus simple des éponges est un tube ou un vase appelé « asconoïde », mais cela limite considérablement la taille de l'animal. La structure corporelle est caractérisée par un spongocèle en forme de tige entouré d'une seule couche de choanocytes. Si elle est simplement agrandie, le rapport entre son volume et sa surface augmente, car la surface augmente comme le carré de la longueur ou de la largeur tandis que le volume augmente proportionnellement au cube. La quantité de tissu qui a besoin de nourriture et d'oxygène est déterminée par le volume, mais la capacité de pompage qui fournit de la nourriture et de l'oxygène dépend de la surface couverte par les choanocytes. Les éponges asconoïdes dépassent rarement 1 mm (0,039 po) de diamètre.
Schéma d'une éponge syconoïde
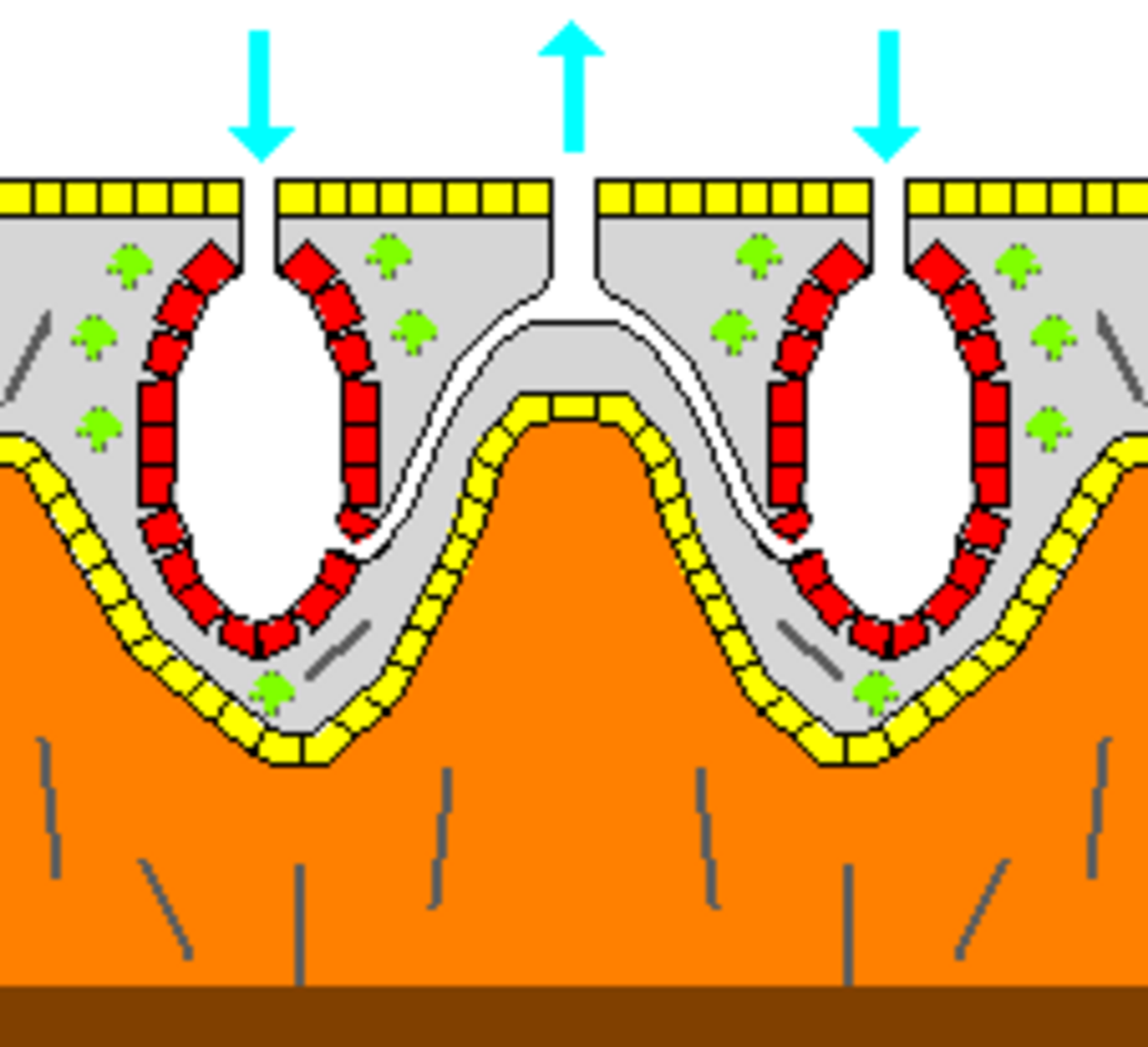
Certaines éponges surmontent cette limitation en adoptant la structure « syconoïde », dans laquelle la paroi du corps est plissée . Les poches intérieures des plis sont tapissées de choanocytes, qui se connectent aux poches extérieures des plis par des ostia. Cette augmentation du nombre de choanocytes et donc de la capacité de pompage permet aux éponges syconoïdes de grandir jusqu'à quelques centimètres de diamètre.
Le motif « leuconoïde » augmente encore la capacité de pompage en remplissant presque entièrement l'intérieur de mésophyle qui contient un réseau de chambres tapissées de choanocytes et reliées entre elles et aux entrées et sorties d'eau par des tubes. Les éponges leuconides atteignent plus d'un mètre de diamètre et le fait que la croissance dans n'importe quelle direction augmente le nombre de chambres de choanocytes leur permet de prendre une plus large gamme de formes, par exemple, des éponges « encroûtantes » dont les formes suivent celles des surfaces auxquelles elles s'attachent. Toutes les éponges d'eau douce et la plupart des éponges marines d'eau peu profonde ont des corps leuconides. Les réseaux de passages d'eau dans les éponges de verre sont similaires à la structure leuconide.
Dans les trois types de structures, la section transversale des régions tapissées de choanocytes est bien plus grande que celle des canaux d'admission et de sortie. Cela ralentit le flux à proximité des choanocytes et leur permet ainsi de piéger plus facilement les particules alimentaires. Par exemple, chez Leuconia , une petite éponge leuconoïde d'environ 10 centimètres (3,9 pouces) de haut et 1 centimètre (0,39 pouce) de diamètre, l'eau pénètre dans chacun des plus de 80 000 canaux d'admission à 6 cm par minute . Cependant, comme Leuconia possède plus de 2 millions de chambres flagellées dont le diamètre combiné est bien supérieur à celui des canaux, le flux d'eau à travers les chambres ralentit à 3,6 cm par heure , ce qui permet aux choanocytes de capturer facilement la nourriture. Toute l'eau est expulsée par un seul osculum à environ 8,5 cm par seconde , suffisamment rapidement pour transporter les déchets à une certaine distance.
En zoologie, un squelette est toute structure relativement rigide d'un animal, qu'elle soit dotée d'articulations ou non et qu'elle soit biominéralisée ou non . Le mésohyle fonctionne comme un endosquelette dans la plupart des éponges et est le seul squelette des éponges molles qui encroûtent des surfaces dures comme les rochers. Le plus souvent, le mésohyle est rigidifié par des spicules minéraux , par des fibres de spongine ou les deux. Les spicules, qui sont présents dans la plupart des espèces mais pas dans toutes, peuvent être constitués de silice ou de carbonate de calcium et varient en forme de simples tiges à des « étoiles » tridimensionnelles comportant jusqu'à six rayons. Les spicules sont produits par des cellules sclérocytes , et peuvent être séparés, reliés par des articulations ou fusionnés.
Certaines éponges sécrètent également des exosquelettes qui se trouvent complètement à l'extérieur de leurs composants organiques. Par exemple, les sclérosponges (« éponges dures ») ont des exosquelettes massifs en carbonate de calcium sur lesquels la matière organique forme une fine couche avec des chambres à choanocytes dans des puits du minéral. Ces exosquelettes sont sécrétés par les pinacocytes qui forment la peau des animaux.
Fonctions vitales
Spongia officinalis , « l'éponge de cuisine », est gris foncé lorsqu'elle est vivante.
Mouvement
Bien que les éponges adultes soient fondamentalement des animaux sessiles , certaines espèces marines et d'eau douce peuvent se déplacer sur le fond marin à une vitesse de 1 à 4 mm (0,039 à 0,157 po) par jour, grâce aux mouvements des pinacocytes et d'autres cellules, semblables à ceux des amibes . Quelques espèces peuvent contracter tout leur corps et beaucoup peuvent fermer leurs oscules et leurs ostiums. Les juvéniles dérivent ou nagent librement, tandis que les adultes sont stationnaires.
Les éponges n'ont pas de systèmes circulatoire , respiratoire , digestif et excréteur distincts - à la place, le système d'écoulement de l'eau prend en charge toutes ces fonctions. Elles filtrent les particules alimentaires de l'eau qui les traverse. Les particules de plus de 50 micromètres ne peuvent pas pénétrer dans les ostia et les pinacocytes les consomment par phagocytose (engloutissement et digestion intracellulaire). Les particules de 0,5 μm à 50 μm sont piégées dans les ostia, qui se rétrécissent des extrémités extérieures vers les extrémités intérieures. Ces particules sont consommées par les pinacocytes ou par les archéocytes qui s'extrudent partiellement à travers les parois des ostia. Les particules de la taille d'une bactérie, inférieures à 0,5 micromètre, traversent les ostia et sont capturées et consommées par les choanocytes . Étant donné que les plus petites particules sont de loin les plus courantes, les choanocytes capturent généralement 80 % de l'approvisionnement en nourriture d'une éponge. Les archéocytes transportent la nourriture emballée dans des vésicules depuis les cellules qui digèrent directement la nourriture vers celles qui ne le font pas. Au moins une espèce d'éponge possède des fibres internes qui fonctionnent comme des pistes utilisées par les archéocytes transporteurs de nutriments, et ces pistes déplacent également des objets inertes.
On prétendait autrefois que les éponges de verre pouvaient vivre de nutriments dissous dans l'eau de mer et étaient très opposées au limon. Cependant, une étude de 2007 n'a trouvé aucune preuve de cela et a conclu qu'elles extraient les bactéries et autres micro-organismes de l'eau de manière très efficace (environ 79 %) et traitent les grains de sédiments en suspension pour extraire ces proies. Les corps du collier digèrent la nourriture et la distribuent enveloppée dans des vésicules qui sont transportées par des molécules « motrices » de dynéine le long de faisceaux de microtubules qui parcourent le syncytium .
Les cellules des éponges absorbent l'oxygène par diffusion de l'eau dans les cellules lorsque l'eau s'écoule dans le corps, dans lequel le dioxyde de carbone et d'autres déchets solubles tels que l'ammoniac se diffusent également. Les archéocytes éliminent les particules minérales qui menacent de bloquer les ostia, les transportent à travers le mésohyle et les déversent généralement dans le courant d'eau sortant, bien que certaines espèces les incorporent dans leur squelette.
Les éponges carnivores
L'éponge carnivore arbre de ping-pong, Chondrocladia lampadiglobus
Dans les eaux où l'approvisionnement en particules alimentaires est très faible, certaines espèces se nourrissent de crustacés et d'autres petits animaux. Jusqu'à présent, seules 137 espèces ont été découvertes. La plupart appartiennent à la famille des Cladorhizidae , mais quelques membres des Guitarridae et des Esperiopsidae sont également carnivores. Dans la plupart des cas, on sait peu de choses sur la manière dont ils capturent réellement leurs proies, bien que l'on pense que certaines espèces utilisent soit des fils collants, soit des spicules crochus . La plupart des éponges carnivores vivent dans des eaux profondes, jusqu'à 8 840 m (5,49 mi), et le développement de techniques d'exploration des grands fonds devrait conduire à la découverte de plusieurs autres. Cependant, une espèce a été trouvée dans des grottes méditerranéennes à des profondeurs de 17 à 23 m (56 à 75 pieds), à côté des éponges filtreuses plus habituelles . Les prédateurs vivant dans les grottes capturent des crustacés de moins de 1 mm (0,039 po) de long en les emmêlant avec des fils fins, les digèrent en les enveloppant de fils supplémentaires au cours de quelques jours, puis reviennent à leur forme normale ; il n'y a aucune preuve qu'ils utilisent du venin .
La plupart des éponges carnivores connues ont complètement perdu leur système d'écoulement de l'eau et leurs choanocytes . Cependant, le genre Chondrocladia utilise un système d'écoulement de l'eau hautement modifié pour gonfler des structures en forme de ballon qui sont utilisées pour capturer des proies.
Endosymbiotes
Les éponges d'eau douce hébergent souvent des algues vertes comme endosymbiotes au sein des archéocytes et d'autres cellules et bénéficient des nutriments produits par les algues. De nombreuses espèces marines hébergent d'autres organismes photosynthétiques , le plus souvent des cyanobactéries mais dans certains cas des dinoflagellés . Les cyanobactéries symbiotiques peuvent constituer un tiers de la masse totale des tissus vivants de certaines éponges, et certaines éponges tirent 48 à 80 % de leur approvisionnement en énergie de ces micro-organismes. En 2008, une équipe de l'Université de Stuttgart a signalé que des spicules en silice conduisent la lumière dans le mésohyle , où vivent les endosymbiotes photosynthétiques. Les éponges qui hébergent des organismes photosynthétiques sont plus courantes dans les eaux relativement pauvres en particules alimentaires et ont souvent des formes de feuilles qui maximisent la quantité de lumière solaire qu'elles collectent.
Les éponges ne possèdent pas le système immunitaire complexe de la plupart des autres animaux. Cependant, elles rejettent les greffes d'autres espèces mais les acceptent d'autres membres de leur propre espèce. Chez quelques espèces marines, les cellules grises jouent le rôle principal dans le rejet de matériel étranger. Lorsqu'elles sont envahies, elles produisent une substance chimique qui arrête le mouvement des autres cellules dans la zone affectée, empêchant ainsi l'intrus d'utiliser les systèmes de transport internes de l'éponge. Si l'intrusion persiste, les cellules grises se concentrent dans la zone et libèrent des toxines qui tuent toutes les cellules de la zone. Le système « immunitaire » peut rester dans cet état activé jusqu'à trois semaines.
Les éponges ont trois méthodes de reproduction asexuée : après fragmentation, par bourgeonnement et en produisant des gemmules . Les fragments d'éponges peuvent être détachés par les courants ou les vagues. Elles utilisent la mobilité de leurs pinacocytes et de leurs choanocytes et le remodelage du mésohyle pour se rattacher à une surface appropriée, puis se reconstruire en tant qu'éponges petites mais fonctionnelles au cours de plusieurs jours. Les mêmes capacités permettent aux éponges qui ont été pressées à travers un tissu fin de se régénérer. Un fragment d'éponge ne peut se régénérer que s'il contient à la fois des collencytes pour produire du mésohyle et des archéocytes pour produire tous les autres types de cellules. Très peu d'espèces se reproduisent par bourgeonnement.
Les gemmules sont des « gousses de survie » que quelques éponges marines et de nombreuses espèces d'eau douce produisent par milliers lorsqu'elles meurent et que certaines, principalement des espèces d'eau douce, produisent régulièrement en automne. Les spongocytes fabriquent des gemmules en enveloppant des coquilles de spongine, souvent renforcées par des spicules, des amas ronds d' archéocytes remplis de nutriments. Les gemmules d'eau douce peuvent également inclure des symbiotes photosynthétiques. Les gemmules deviennent alors dormantes et, dans cet état, peuvent survivre au froid, à la sécheresse, au manque d'oxygène et aux variations extrêmes de salinité . Les gemmules d'eau douce ne reprennent souvent vie que lorsque la température baisse, reste froide pendant quelques mois puis atteint un niveau presque « normal ». Lorsqu'une gemmule germe, les archéocytes situés autour de l'extérieur de l'amas se transforment en pinacocytes , une membrane recouvrant un pore de la coquille éclate, l'amas de cellules émerge lentement et la plupart des archéocytes restants se transforment en d'autres types de cellules nécessaires à la fabrication d'une éponge fonctionnelle. Les gemmules de la même espèce mais d'individus différents peuvent unir leurs forces pour former une éponge. Certaines gemmules sont retenues dans l'éponge mère et, au printemps, il peut être difficile de dire si une vieille éponge a repris vie ou a été « recolonisée » par ses propres gemmules.
Sexuel
La plupart des éponges sont hermaphrodites (fonctionnent simultanément comme les deux sexes), bien que les éponges n'aient pas de gonades (organes reproducteurs). Les spermatozoïdes sont produits par des choanocytes ou des chambres de choanocytes entières qui s'enfoncent dans le mésohyle et forment des kystes spermatiques tandis que les œufs sont formés par la transformation d' archéocytes , ou de choanocytes chez certaines espèces. Chaque œuf acquiert généralement un jaune en consommant des « cellules nourricières ». Pendant la ponte, les spermatozoïdes jaillissent de leurs kystes et sont expulsés via l' oscule . S'ils entrent en contact avec une autre éponge de la même espèce, le flux d'eau les transporte vers les choanocytes qui les engloutissent mais, au lieu de les digérer, se métamorphosent en une forme amiboïde et transportent le sperme à travers le mésohyle jusqu'aux œufs, qui dans la plupart des cas engloutissent le porteur et sa cargaison.
Quelques espèces libèrent des œufs fécondés dans l'eau, mais la plupart les conservent jusqu'à leur éclosion. En conservant les œufs, les parents peuvent transférer des micro-organismes symbiotiques directement à leur progéniture par transmission verticale , tandis que les espèces qui libèrent leurs œufs dans l'eau doivent acquérir des symbiotes horizontalement (une combinaison des deux est probablement la plus courante, où les larves avec des symbiotes transmis verticalement en acquièrent également d'autres horizontalement). Il existe quatre types de larves, mais toutes sont des boules de cellules lécithotrophes (non nourrissantes) avec une couche externe de cellules dont les flagelles ou les cils permettent aux larves de se déplacer. Après avoir nagé pendant quelques jours, les larves coulent et rampent jusqu'à ce qu'elles trouvent un endroit où s'installer. La plupart des cellules se transforment en archéocytes, puis en types appropriés à leur emplacement dans une éponge adulte miniature.
Les embryons d'éponges de verre commencent par se diviser en cellules séparées, mais une fois que 32 cellules se sont formées, ils se transforment rapidement en larves qui sont extérieurement ovoïdes avec une bande de cils autour du milieu qu'elles utilisent pour se déplacer, mais qui ont intérieurement la structure typique des éponges de verre de spicules avec un syncitium principal en forme de toile d'araignée drapé autour et entre eux et des choanosyncyties avec de multiples corps en collier au centre. Les larves quittent ensuite le corps de leurs parents.
Méiose
La progression cytologique de l'oogénèse et de la spermatogenèse ( gamétogenèse ) des porifères est très similaire à celle des autres métazoaires. La plupart des gènes de l'ensemble classique des gènes méiotiques , y compris les gènes de recombinaison de l'ADN et de réparation des cassures double brin, qui sont conservés chez les eucaryotes , sont exprimés dans les éponges (par exemple Geodia hentscheli et Geodia phlegraei ). Étant donné que les porifères sont considérés comme les premiers animaux divergents, ces résultats indiquent que la boîte à outils de base de la méiose, y compris les capacités de recombinaison et de réparation de l'ADN, était présente au début de l'évolution des eucaryotes.
Cycle de vie
Les éponges des régions tempérées vivent au plus quelques années, mais certaines espèces tropicales et peut-être certaines espèces des profondeurs océaniques peuvent vivre 200 ans ou plus. Certaines démosponges calcifiées ne grandissent que de 0,2 mm (0,0079 po) par an et, si ce taux est constant, les spécimens de 1 m (3,3 pi) de large doivent avoir environ 5 000 ans. Certaines éponges commencent la reproduction sexuée à l'âge de quelques semaines seulement, tandis que d'autres attendent d'avoir plusieurs années.
Coordination des activités
Les éponges adultes n'ont pas de neurones ni de tissu nerveux . Cependant, la plupart des espèces ont la capacité d'effectuer des mouvements coordonnés dans tout leur corps, principalement des contractions des pinacocytes , qui compriment les canaux d'eau et expulsent ainsi l'excès de sédiments et d'autres substances qui peuvent provoquer des blocages. Certaines espèces peuvent contracter l' oscule indépendamment du reste du corps. Les éponges peuvent également se contracter afin de réduire la zone vulnérable aux attaques des prédateurs. Dans les cas où deux éponges sont fusionnées, par exemple s'il y a un gros bourgeon non séparé, ces ondes de contraction deviennent lentement coordonnées dans les deux « jumeaux siamois ». Le mécanisme de coordination est inconnu, mais pourrait impliquer des produits chimiques similaires aux neurotransmetteurs . Cependant, les éponges de verre transmettent rapidement des impulsions électriques à travers toutes les parties du syncytium et utilisent cela pour arrêter le mouvement de leurs flagelles si l'eau entrante contient des toxines ou des sédiments excessifs. les myocytes sont responsables de la fermeture de l’oscule et de la transmission des signaux entre les différentes parties du corps.
Les éponges contiennent des gènes très similaires à ceux qui contiennent la « recette » de la densité post- synaptique , une importante structure de réception de signaux dans les neurones de tous les autres animaux. Cependant, chez les éponges, ces gènes ne sont activés que dans les « cellules en flacon » qui n'apparaissent que chez les larves et peuvent fournir une certaine capacité sensorielle pendant que les larves nagent. Cela soulève des questions quant à savoir si les cellules en flacon représentent les prédécesseurs de vrais neurones ou sont la preuve que les ancêtres des éponges avaient de vrais neurones mais les ont perdus en s'adaptant à un mode de vie sessile.
Les éponges sont réparties dans le monde entier et vivent dans une grande variété d'habitats océaniques, des régions polaires aux tropiques. La plupart vivent dans des eaux calmes et claires, car les sédiments soulevés par les vagues ou les courants obstrueraient leurs pores, ce qui les empêcherait de se nourrir et de respirer. Le plus grand nombre d'éponges se trouvent généralement sur des surfaces fermes telles que les rochers, mais certaines éponges peuvent se fixer sur des sédiments mous au moyen d'une base semblable à une racine.
Les éponges sont plus abondantes mais moins diversifiées dans les eaux tempérées que dans les eaux tropicales, peut-être parce que les organismes qui se nourrissent d'éponges sont plus abondants dans les eaux tropicales. Les éponges de verre sont les plus courantes dans les eaux polaires et dans les profondeurs des mers tempérées et tropicales, car leur construction très poreuse leur permet d'extraire de la nourriture de ces eaux pauvres en ressources avec un minimum d'effort. Les démosponges et les éponges calcaires sont abondantes et diversifiées dans les eaux non polaires moins profondes.
Les différentes classes d'éponges vivent dans des habitats différents :
En tant que producteurs primaires
Les éponges dotées d' endosymbiotes photosynthétiques produisent jusqu'à trois fois plus d'oxygène qu'elles n'en consomment, ainsi que plus de matière organique qu'elles n'en consomment. Ces contributions aux ressources de leurs habitats sont importantes le long de la Grande Barrière de corail australienne, mais relativement mineures dans les Caraïbes.
Défenses
Trous faits par une éponge clionaid (produisant la trace Entobia ) après la mort d'une coquille bivalve moderne de l'espèce Mercenaria mercenaria , de Caroline du NordGros plan de l'éponge perforante Entobia dans une valve d'huître moderne. Notez les chambres reliées par de courts tunnels.
De nombreuses éponges perdent des spicules , formant un tapis dense de plusieurs mètres de profondeur qui éloigne les échinodermes qui, autrement, les attaqueraient. Elles produisent également des toxines qui empêchent d'autres organismes sessiles tels que les bryozoaires ou les ascidies de pousser sur ou à proximité d'elles, ce qui fait des éponges des compétiteurs très efficaces pour l'espace vital. L'un des nombreux exemples est l'agéliférine .
Quelques espèces, comme l' éponge de feu des Caraïbes Tedania ignis , provoquent une éruption cutanée sévère chez les humains qui les manipulent. Les tortues et certains poissons se nourrissent principalement d'éponges. On dit souvent que les éponges produisent des défenses chimiques contre ces prédateurs. Cependant, les expériences n'ont pas permis d'établir une relation entre la toxicité des produits chimiques produits par les éponges et leur goût pour les poissons, ce qui diminuerait l'utilité des défenses chimiques comme moyen de dissuasion. La prédation par les poissons peut même aider à propager les éponges en détachant des fragments. Cependant, certaines études ont montré que les poissons montraient une préférence pour les éponges non protégées chimiquement, et une autre étude a constaté que des niveaux élevés de prédation des coraux prédisaient la présence d'espèces protégées chimiquement.
Les éponges de verre ne produisent aucun produit chimique toxique et vivent dans des eaux très profondes où les prédateurs sont rares.
Prédation
Les mouches à éponge, également appelées mouches à éponge ( Neuroptera , Sisyridae ), sont des prédateurs spécialisés dans les éponges d'eau douce. La femelle pond ses œufs sur la végétation surplombant l'eau. Les larves éclosent et tombent dans l'eau où elles recherchent des éponges pour se nourrir. Elles utilisent leurs pièces buccales allongées pour percer l'éponge et aspirer les fluides qu'elle contient. Les larves de certaines espèces s'accrochent à la surface de l'éponge tandis que d'autres se réfugient dans les cavités internes de l'éponge. Les larves adultes quittent l'eau et tissent un cocon dans lequel se nymphoser.
Bioérosion
L'éponge à foie de poulet des Caraïbes Chondrilla nucula sécrète des toxines qui tuent les polypes coralliens , permettant aux éponges de se développer sur les squelettes coralliens. D'autres, en particulier dans la famille des Clionaidae , utilisent des substances corrosives sécrétées par leurs archéocytes pour creuser des tunnels dans les roches, les coraux et les coquilles de mollusques morts . Les éponges peuvent retirer jusqu'à 1 m (3,3 pieds) par an des récifs, créant des entailles visibles juste en dessous du niveau de la marée basse.
Maladies
Les éponges des Caraïbes du genre Aplysina souffrent du syndrome des bandes rouges d'Aplysina. Ce syndrome provoque le développement d'une ou plusieurs bandes de couleur rouille, parfois avec des bandes adjacentes de tissu nécrotique . Ces lésions peuvent encercler complètement les branches de l'éponge. La maladie semble être contagieuse et touche environ dix pour cent des A. cauliformis des récifs des Bahamas. Les bandes de couleur rouille sont causées par une cyanobactérie , mais on ne sait pas si cet organisme est réellement à l'origine de la maladie.
Collaboration avec d’autres organismes
En plus d'héberger des endosymbiotes photosynthétiques, les éponges sont connues pour leur large gamme de collaborations avec d'autres organismes. L'éponge encroûtante relativement grande Lissodendoryx colombiensis est plus commune sur les surfaces rocheuses, mais a étendu son aire de répartition aux prairies d'herbiers marins en se laissant encercler ou envahir par des éponges d'herbiers marins, qui sont désagréables pour les étoiles de mer locales et protègent donc Lissodendoryx contre elles ; en retour, les éponges d'herbiers marins obtiennent des positions plus élevées, loin des sédiments du fond marin.
Les crevettes du genre Synalpheus forment des colonies dans des éponges, et chaque espèce de crevette habite une espèce d'éponge différente, ce qui fait de Synalpheus l'un des genres de crustacés les plus diversifiés . Plus précisément, Synalpheus regalis utilise l'éponge non seulement comme source de nourriture, mais aussi comme moyen de défense contre d'autres crevettes et prédateurs. Jusqu'à 16 000 individus habitent une seule éponge caouanne , se nourrissant des particules plus grosses qui s'accumulent sur l'éponge lorsqu'elle filtre l'océan pour se nourrir. D'autres crustacés tels que les bernard-l'ermite ont généralement une espèce spécifique d'éponge, Pseudospongosorites , qui pousse sur eux car l'éponge et le crabe occupent tous deux des coquilles de gastéropodes jusqu'à ce que le crabe et l'éponge dépassent la coquille, ce qui fait que le crabe utilise finalement le corps de l'éponge comme protection au lieu de la coquille jusqu'à ce que le crabe trouve une coquille de remplacement appropriée.
Aire de répartition bathymétrique de certaines espèces d'éponges. Demosponge Samus anonymus (jusqu'à 50 m), hexaactinellidé Scleroplegma lanterna (~ 100-600 m), hexaactinellidé Aulocalyx irrégulières (~ 550-915 m), démosponge lithistidé Neoaulaxinia persicum (~ 500-1700 m)Réseau alimentaire généralisé pour les récifs d'éponges
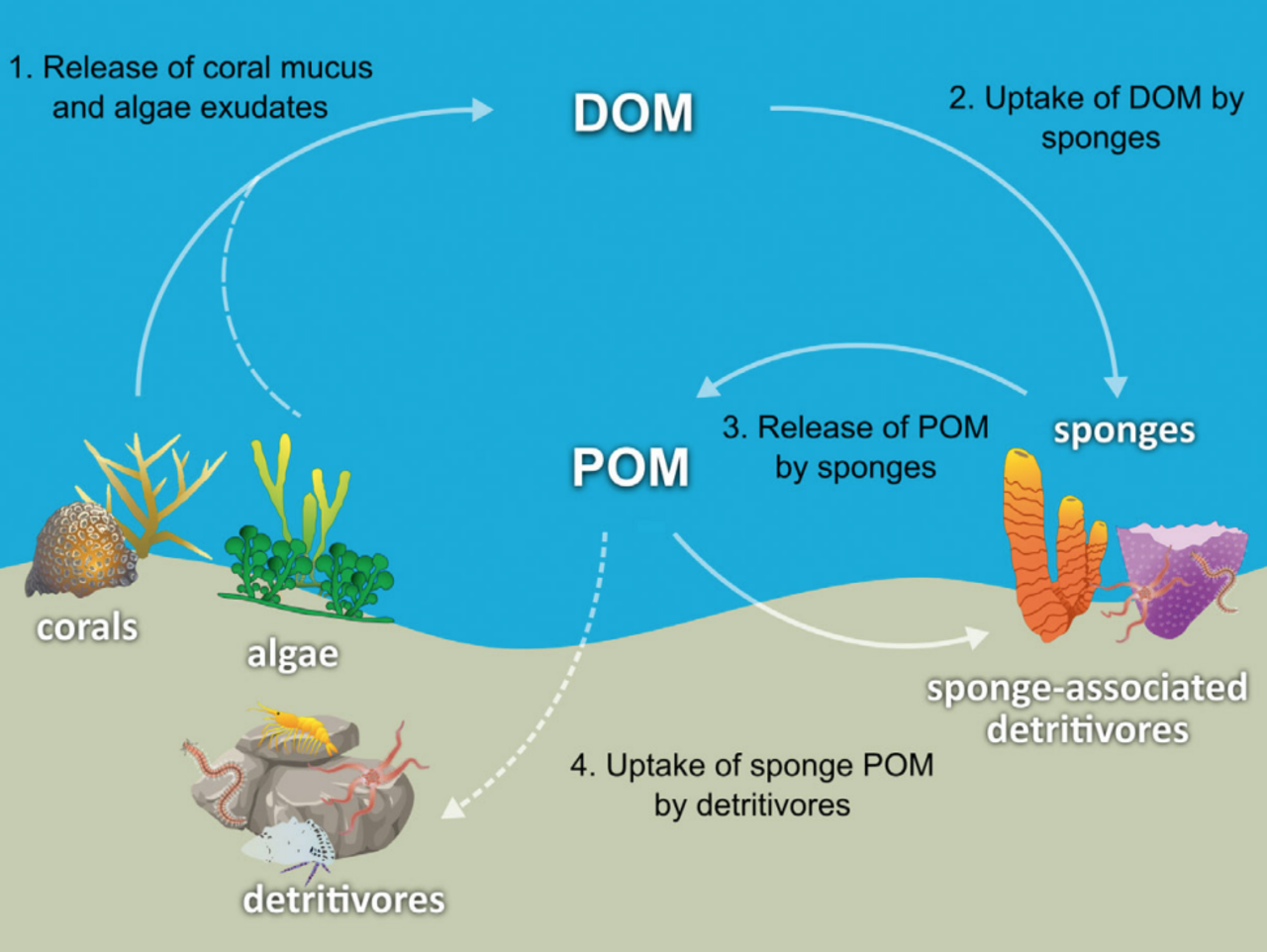
L'hypothèse a été émise que les éponges des récifs coralliens facilitent le transfert de la matière organique dérivée du corail à leurs détritivores associés via la production de détritus d'éponge, comme le montre le diagramme. Plusieurs espèces d'éponges sont capables de convertir la matière organique dérivée du corail en détritus d'éponge, et de transférer la matière organique produite par les coraux plus haut dans le réseau trophique récifal. Les coraux libèrent de la matière organique sous forme de mucus dissous et particulaire, ainsi que de matière cellulaire telle que le Symbiodinium expulsé .
La matière organique pourrait être transférée des coraux aux éponges par toutes ces voies, mais la DOM constitue probablement la plus grande fraction, car la majorité (56 à 80 %) du mucus corallien se dissout dans la colonne d'eau, et la perte de carbone fixe due à l'expulsion de Symbiodinium est généralement négligeable (0,01 %) par rapport à la libération de mucus (jusqu'à environ 40 %). La matière organique dérivée des coraux pourrait également être transférée indirectement aux éponges par l'intermédiaire de bactéries, qui peuvent également consommer du mucus corallien.
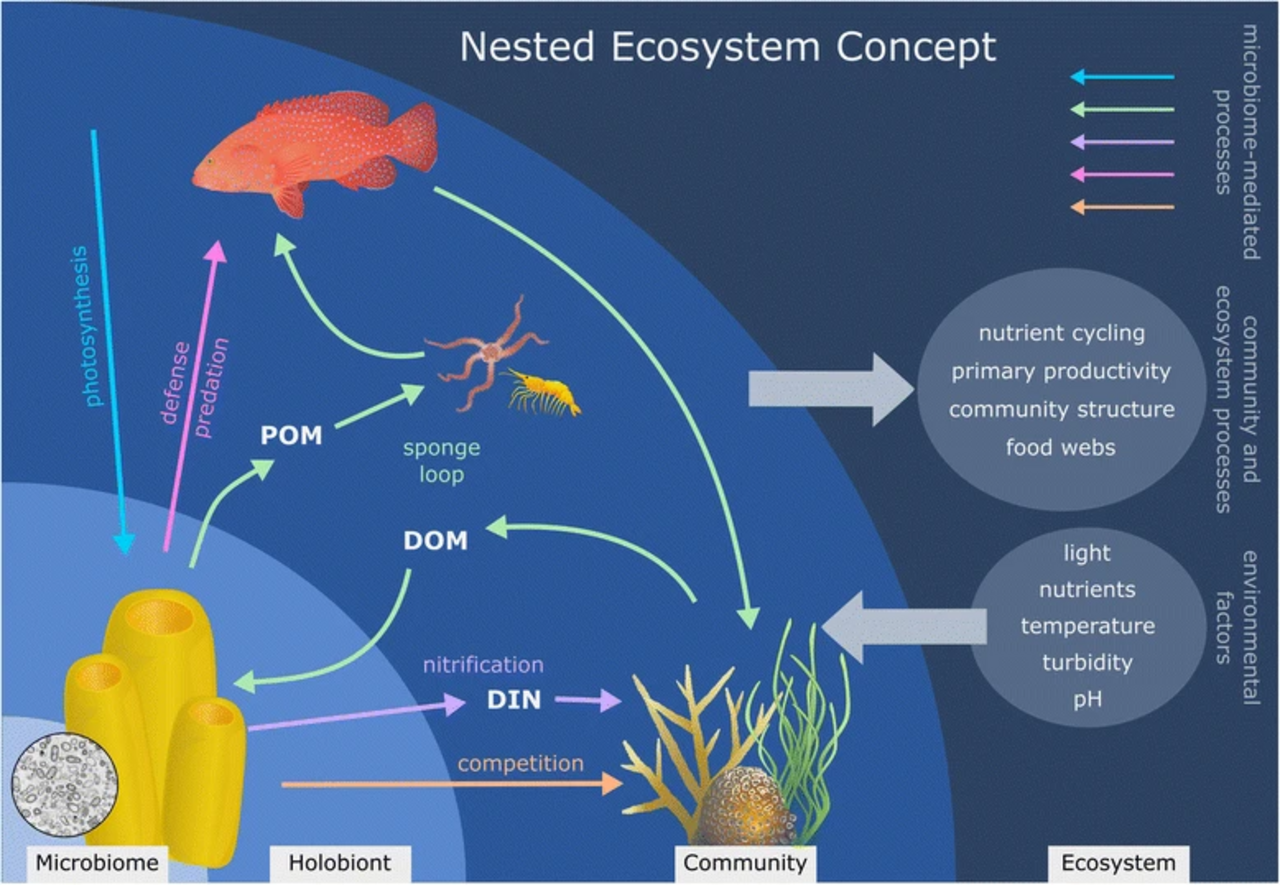
Hypothèse de la boucle de l'éponge. Étapes de la voie de la boucle de l'éponge : (1) les coraux et les algues libèrent des exsudats sous forme de matière organique dissoute (DOM), (2) les éponges absorbent la DOM, (3) les éponges libèrent de la matière organique particulaire détritique (POM), (4) les détritus d'éponge (POM) sont absorbés par les détritivores associés aux éponges et les détritivores libres . L'holobionte spongieux. L' holobionte spongieux est un exemple du concept d'écosystèmes imbriqués. Les fonctions clés exercées par le microbiome (flèches colorées) influencent le fonctionnement de l'holobionte et, par des effets en cascade, influencent ensuite la structure de la communauté et le fonctionnement de l'écosystème. Les facteurs environnementaux agissent à plusieurs échelles pour modifier les processus à l'échelle du microbiome, de l'holobionte, de la communauté et de l'écosystème. Ainsi, les facteurs qui altèrent le fonctionnement du microbiome peuvent entraîner des changements au niveau de l'holobionte, de la communauté ou même de l'écosystème et vice versa, illustrant la nécessité de prendre en compte plusieurs échelles lors de l'évaluation du fonctionnement dans les écosystèmes imbriqués. (DOM : matière organique dissoute , POM : matière organique particulaire , DIN : azote inorganique dissous)
Holobionte éponge
Outre une relation symbiotique un à un , il est possible pour un hôte de devenir symbiotique avec un consortium microbien , ce qui donne naissance à un microbiome d'éponge diversifié . Les éponges sont capables d'héberger une large gamme de communautés microbiennes qui peuvent également être très spécifiques. Les communautés microbiennes qui forment une relation symbiotique avec l'éponge peuvent représenter jusqu'à 35 % de la biomasse de son hôte. Le terme pour cette relation symbiotique spécifique, où un consortium microbien s'associe à un hôte, est appelé relation holobiotique . L'éponge ainsi que la communauté microbienne qui lui est associée produiront une large gamme de métabolites secondaires qui aident à la protéger contre les prédateurs par des mécanismes tels que la défense chimique .
Certaines de ces relations incluent des endosymbiotes au sein des cellules bactériocytes et des cyanobactéries ou microalgues trouvées sous la couche cellulaire du pinacoderme où elles peuvent recevoir la plus grande quantité de lumière, utilisée pour la phototrophie. Elles peuvent héberger plus de 50 phylums microbiens différents et phylums candidats, notamment les Alphaprotoebactéries, les Actinomycetota , les Chloroflexota , les Nitrospirota , les « Cyanobactéries », les taxons Gamma-, le phylum candidat Poribacteria et les Thaumarchaea .
Systématique
Taxonomie
Carl Linnaeus , qui a classé la plupart des espèces d'animaux sessiles comme appartenant à l'ordre des Zoophytes dans la classe des Vermes , a identifié par erreur le genre Spongia comme des plantes de l'ordre des algues . Pendant longtemps par la suite, les éponges ont été assignées au sous-royaume des Parazoa (« à côté des animaux ») séparé des Eumetazoa qui formaient le reste du royaume Animalia .
Le phylum Porifera est en outre divisé en classes principalement en fonction de la composition de leurs squelettes :
Les cellules Calcarea ont un squelette fait de calcite , une forme de carbonate de calcium , qui peut former des spicules séparés ou de grandes masses. Toutes les cellules ont un seul noyau et une seule membrane.
La plupart des démosponges ont des spicules de silicate ou des fibres de spongine ou les deux dans leurs tissus mous. Cependant, quelques-unes ont également des squelettes externes massifs faits d' aragonite , une autre forme de carbonate de calcium. Toutes les cellules ont un seul noyau et une seule membrane.
Dans les années 1970, les éponges à squelette massif de carbonate de calcium ont été classées dans une classe distincte, les Sclerospongiae , autrement appelées « éponges coralliennes ». Cependant, dans les années 1980, on a découvert qu'elles appartenaient toutes soit aux Calcarea, soit aux Demospongiae.
Jusqu'à présent, les publications scientifiques ont identifié environ 9 000 espèces de porifères, dont environ 400 sont des éponges de verre, environ 500 sont des espèces calcaires et le reste sont des démosponges. Cependant, certains types d'habitats, les parois rocheuses verticales et les grottes et les galeries dans les rochers et les blocs de corail, ont été très peu étudiés, même dans les mers peu profondes.
Cours
Les éponges étaient traditionnellement réparties en trois classes : les éponges calcaires (Calcarea), les éponges de verre (Hexactinellida) et les démosponges (Demospongiae). Cependant, des études ont maintenant montré que les Homoscleromorpha , un groupe qui appartiendrait aux Demospongiae , ont une relation génétique bien distincte des autres classes d'éponges. Par conséquent, elles ont récemment été reconnues comme la quatrième classe d'éponges.
Les éponges sont divisées en classes principalement en fonction de la composition de leur squelette : Celles-ci sont classées par ordre évolutif comme indiqué ci-dessous dans l'ordre croissant de leur évolution de haut en bas :
Phylogénie
La phylogénie des éponges a été largement débattue depuis l'avènement de la phylogénétique . Considérée à l'origine comme le phylum animal le plus basal, il existe désormais de nombreuses preuves que Ctenophora pourrait détenir ce titre. De plus, la monophylie du phylum est désormais remise en question. Plusieurs études ont conclu que tous les autres animaux sont issus des éponges et constatent généralement que les éponges calcaires et les Homoscleromorpha sont plus proches des autres animaux que des démosponges . Les relations internes des Porifera se sont avérées moins incertaines. Une relation étroite entre Homoscleromorpha et Calcarea a été retrouvée dans presque toutes les études, qu'elles soutiennent ou non la monophylie des éponges ou des eumétazoaires. La position des éponges de verre est également assez certaine, la majorité des études les retrouvant comme sœurs des démosponges. Ainsi, l’incertitude à la base de l’arbre généalogique des animaux est probablement mieux représentée par le cladogramme ci-dessous.
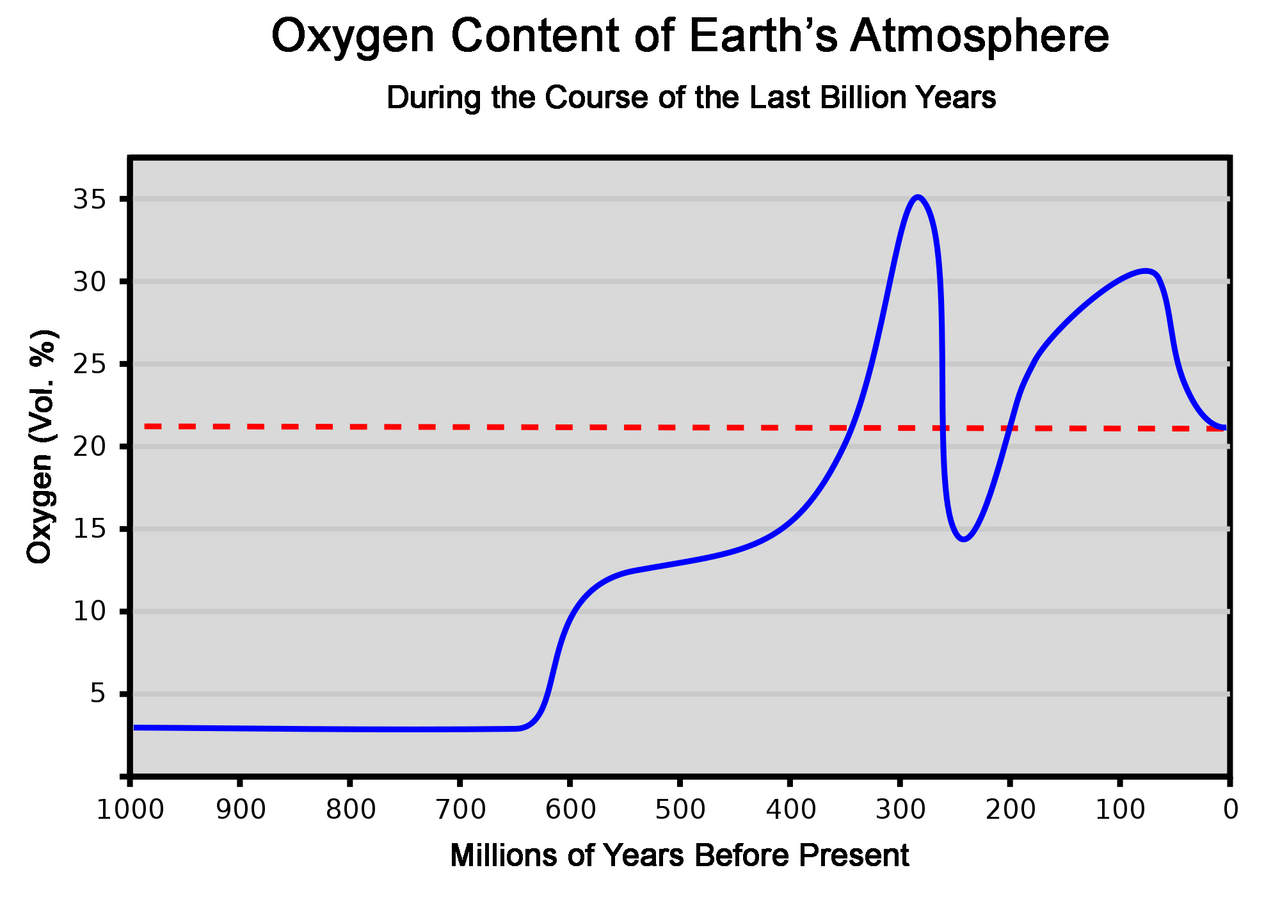
Français Bien que les horloges moléculaires et les biomarqueurs suggèrent que les éponges existaient bien avant l' explosion de la vie au Cambrien, les spicules de silice comme ceux des démosponges sont absents du registre fossile jusqu'au Cambrien. Un rapport non corroboré de 2002 existe sur des spicules dans des roches datées d'environ 750 millions d'années . Des éponges fossiles bien préservées datant d'environ 580 millions d'années de la période édiacarienne ont été trouvées dans la formation de Doushantuo . Ces fossiles, qui comprennent : des spicules ; des pinacocytes ; des porocytes ; des archéocytes ; des sclérocytes ; et la cavité interne, ont été classés comme des démosponges. Des fossiles d' éponges de verre ont été trouvés il y a environ 540 millions d'années dans des roches en Australie, en Chine et en Mongolie. Des éponges du Cambrien précoce du Mexique appartenant au genre Kiwetinokia montrent des preuves de la fusion de plusieurs spicules plus petits pour former un seul grand spicule. Des spicules de carbonate de calcium d'éponges calcaires ont été trouvés dans des roches du Cambrien précoce datant d'environ 530 à 523 millions d'années en Australie. D'autres démosponges probables ont été trouvées dans la faune du Cambrien précoce de Chengjiang , il y a 525 à 520 millions d'années . Des fossiles trouvés dans les Territoires du Nord-Ouest canadiens datant d'il y a 890 millions d'années pourraient être des éponges ; si cette découverte est confirmée, elle suggère que les premiers animaux sont apparus avant l'événement d'oxygénation du Néoprotérozoïque.
Teneur en oxygène de l'atmosphère au cours du dernier milliard d'années. Si elle est confirmée, la découverte d'éponges fossilisées datant d'il y a 890 millions d'années serait antérieure à l'événement d'oxygénation du Néoprotérozoïque.
Les éponges d'eau douce semblent être beaucoup plus jeunes, car les premiers fossiles connus datent de la période de l' Éocène moyen, il y a environ 48 à 40 millions d'années . Bien qu'environ 90 % des éponges modernes soient des démosponges , les restes fossilisés de ce type sont moins courants que ceux des autres types, car leurs squelettes sont composés d'éponges relativement molles qui ne se fossilisent pas bien. Les premiers symbiotes d'éponges sont connus depuis le début du Silurien .
Le traceur chimique utilisé est le 24-isopropyl cholestane , un dérivé stable du 24-isopropyl cholestérol , qui serait produit par les démosponges mais pas par les eumétazoaires (« vrais animaux », c'est-à-dire les cnidaires et les bilatériens ). Les choanoflagellés étant considérés comme les plus proches parents unicellulaires des animaux, une équipe de scientifiques a examiné la biochimie et les gènes d'une espèce de choanoflagellés . Ils ont conclu que cette espèce ne pouvait pas produire de 24-isopropyl cholestérol, mais qu'il serait nécessaire d'étudier une gamme plus large de choanoflagellés pour prouver que le 24-isopropyl cholestane fossile ne pouvait être produit que par les démosponges. Bien qu'une publication antérieure ait signalé des traces de 24-isopropyl cholestane dans des roches anciennes datant d' il y a 1 800 millions d'années , des recherches récentes utilisant une série de roches datées beaucoup plus précisément ont révélé que ces biomarqueurs n'apparaissent qu'avant la fin de la glaciation marinoenne , il y a environ 635 millions d'années , et que « l'analyse des biomarqueurs n'a pas encore révélé de preuve convaincante de l'existence d'éponges anciennes antérieures au premier épisode glaciaire néoprotérozoïque d'ampleur mondiale (le Sturtien, il y a environ 713 millions d'années à Oman) ». Bien qu'il ait été avancé que ce « biomarqueur d'éponge » pourrait provenir d'algues marines, des recherches récentes suggèrent que la capacité des algues à produire ce biomarqueur n'a évolué qu'au Carbonifère ; en tant que tel, le biomarqueur reste fortement en faveur de la présence de démosponges au Cryogénien.
Les archéocyathides , que certains classent comme un type d'éponge corallienne, sont des fossiles très courants dans les roches du Cambrien précoce, il y a environ 530 à 520 millions d'années , mais ils ont apparemment disparu à la fin du Cambrien , il y a 490 millions d'années . Il a été suggéré qu'ils ont été produits par : des éponges ; des cnidaires ; des algues ; des foraminifères ; un embranchement d'animaux complètement distinct, les Archaeocyatha ; ou même un règne de vie complètement distinct, appelé Archaeata ou Inferibionta. Depuis les années 1990, les archéocyathides sont considérés comme un groupe distinct d'éponges.
Il est difficile de classer les chancelloriidés dans des classifications d'éponges ou d'animaux plus complexes. Une analyse réalisée en 1996 a conclu qu'ils étaient étroitement liés aux éponges au motif que la structure détaillée des sclérites des chancelloriidés (« plaques d'armure ») est similaire à celle des fibres de spongine, une protéine de collagène , dans les démosponges kératosiques (cornées) modernes telles que Darwinella . Cependant, une autre analyse réalisée en 2002 a conclu que les chancelloriidés ne sont pas des éponges et peuvent être intermédiaires entre les éponges et des animaux plus complexes, entre autres raisons parce que leur peau est plus épaisse et plus étroitement connectée que celle des éponges. En 2008, une analyse détaillée des sclérites des chancelloriidés a conclu qu'ils étaient très similaires à ceux des halkieriidés , des animaux bilatériens mobiles qui ressemblaient à des limaces en cotte de mailles et dont les fossiles se trouvent dans des roches du Cambrien précoce au Cambrien moyen. Si cela est correct, cela créerait un dilemme, car il est extrêmement improbable que des organismes totalement indépendants aient pu développer des sclérites aussi similaires de manière indépendante, mais l'énorme différence dans les structures de leurs corps rend difficile de voir comment ils pourraient être étroitement liés.
Relations avec d’autres groupes d’animaux
Un choanoflagellé
Dans les années 1990, les éponges étaient largement considérées comme un groupe monophylétique , toutes descendant d'un ancêtre commun qui était lui-même une éponge, et comme le « groupe frère » de tous les autres métazoaires (animaux multicellulaires), qui forment eux-mêmes un groupe monophylétique. D'un autre côté, certaines analyses des années 1990 ont également ravivé l'idée que les plus proches parents évolutifs des animaux sont les choanoflagellés , des organismes unicellulaires très similaires aux choanocytes des éponges – ce qui impliquerait que la plupart des métazoaires ont évolué à partir d'ancêtres très semblables aux éponges et donc que les éponges ne sont peut-être pas monophylétiques, car les mêmes ancêtres semblables aux éponges peuvent avoir donné naissance à la fois aux éponges modernes et aux membres non spongieux des métazoaires.
Depuis 2001, des analyses ont conclu que les eumétazoaires (plus complexes que les éponges) sont plus étroitement liés à des groupes particuliers d'éponges qu'à d'autres groupes d'éponges. De telles conclusions impliquent que les éponges ne sont pas monophylétiques, car le dernier ancêtre commun de toutes les éponges serait également un ancêtre direct des eumétazoaires, qui ne sont pas des éponges. Une étude de 2001 basée sur des comparaisons d' ADN ribosomique a conclu que la division la plus fondamentale au sein des éponges se situait entre les éponges de verre et les autres, et que les eumétazoaires étaient plus étroitement liés aux éponges calcaires (celles avec des spicules de carbonate de calcium) qu'aux autres types d'éponges. En 2007, une analyse basée sur des comparaisons d' ARN et une autre basée principalement sur la comparaison de spicules ont conclu que les démosponges et les éponges de verre étaient plus étroitement liées entre elles que l'une ou l'autre ne l'était aux éponges calcaires, qui à leur tour sont plus étroitement liées aux eumétazoaires.
D'autres preuves anatomiques et biochimiques relient les Eumetazoa aux Homoscleromorpha , un sous-groupe de démosponges. Une comparaison réalisée en 2007 de l'ADN nucléaire , excluant les éponges de verre et les méduses à peigne , a conclu que :
les éponges calcaires sont les suivantes les plus proches ;
les autres démosponges sont des « tantes » évolutives de ces groupes ; et
les chancelloriidés , animaux en forme de sac dont les fossiles ont été découverts dans les roches cambriennes , pourraient être des éponges.
Les spermatozoïdes des Homoscleromorpha partagent avec les spermatozoïdes des Eumetazoa des caractéristiques qui leur font défaut. Chez les Homoscleromorpha comme chez les Eumetazoa, les couches de cellules sont liées entre elles par une fixation à une membrane basale en forme de tapis composée principalement de collagène de « type IV » , une forme de collagène que l'on ne trouve pas chez les autres éponges – bien que les fibres de spongine qui renforcent le mésohyle de toutes les démosponges soient similaires au collagène de « type IV ».
Les analyses décrites ci-dessus ont conclu que les éponges sont les ancêtres les plus proches de tous les métazoaires, de tous les animaux multicellulaires, y compris les éponges et les groupes plus complexes. Cependant, une autre comparaison en 2008 de 150 gènes dans chacun des 21 genres, allant des champignons aux humains, mais n'incluant que deux espèces d'éponges, a suggéré que les méduses à peigne ( ctenophora ) sont la lignée la plus basale des métazoaires inclus dans l'échantillon. Si cela est correct, soit les méduses à peigne modernes ont développé leurs structures complexes indépendamment des autres métazoaires, soit les ancêtres des éponges étaient plus complexes et toutes les éponges connues sont des formes considérablement simplifiées. L'étude a recommandé des analyses plus approfondies en utilisant une gamme plus large d'éponges et d'autres métazoaires simples tels que les placozoaires .
Cependant, une nouvelle analyse des données a montré que les algorithmes informatiques utilisés pour l'analyse ont été induits en erreur par la présence de gènes spécifiques de cténophores qui étaient nettement différents de ceux des autres espèces, laissant les éponges soit comme groupe frère de tous les autres animaux, soit comme grade paraphylétique ancestral. Des « arbres généalogiques » construits à l'aide d'une combinaison de toutes les données disponibles - morphologiques, développementales et moléculaires - ont conclu que les éponges sont en fait un groupe monophylétique et qu'avec les cnidaires, elles forment le groupe frère des bilatériens.
Un alignement très important et cohérent de 1 719 protéines à l'échelle des métazoaires, publié en 2017, a montré que (i) les éponges - représentées par Homoscleromorpha, Calcarea, Hexactinellida et Demospongiae - sont monophylétiques, (ii) les éponges sont le groupe frère de tous les autres animaux multicellulaires, (iii) les cténophores émergent comme la deuxième lignée animale la plus ancienne et (iv) les placozoaires émergent comme la troisième lignée animale, suivie par les cnidaires, groupe frère des bilatériens .
En mars 2021, des scientifiques de Dublin ont trouvé des preuves supplémentaires que les éponges sont le groupe frère de tous les autres animaux, tandis qu'en mai 2023, Schultz et al. ont trouvé des modèles de changement irréversible dans la synténie du génome qui fournissent une preuve solide que les cténophores sont plutôt le groupe frère de tous les autres animaux.
Un rapport de 1997 décrit l'utilisation d'éponges comme outil par les grands dauphins de Shark Bay en Australie occidentale. Un dauphin attache une éponge marine à son rostre , qui est probablement utilisée pour le protéger lorsqu'il recherche de la nourriture dans le fond marin sablonneux . Ce comportement, connu sous le nom d'épongement , n'a été observé que dans cette baie et est presque exclusivement observé par les femelles. Une étude de 2005 a conclu que les mères enseignent ce comportement à leurs filles et que tous les utilisateurs d'éponges sont étroitement liés, ce qui suggère qu'il s'agit d'une innovation assez récente.
Les spicules de carbonate de calcium ou de silice de la plupart des genres d'éponges les rendent trop rugueuses pour la plupart des utilisations, mais deux genres, Hippospongia et Spongia , ont des squelettes mous, entièrement fibreux. Les premiers Européens utilisaient des éponges molles à de nombreuses fins, notamment pour le rembourrage des casques, les ustensiles de boisson portables et les filtres à eau municipaux. Jusqu'à l'invention des éponges synthétiques, elles étaient utilisées comme outils de nettoyage, applicateurs de peintures et de glaçures céramiques et contraceptifs discrets . Cependant, au milieu du XXe siècle, la surpêche a conduit les animaux et l'industrie au bord de l'extinction .
L' éponge luffa , également orthographiée loofah , qui est couramment vendue pour être utilisée dans la cuisine ou la douche, n'est pas dérivée d'un animal mais principalement du « squelette » fibreux de la courge éponge ( Luffa aegyptiaca , Cucurbitacées ).
Composés antibiotiques
Les éponges ont un potentiel médicinal en raison de la présence dans les éponges elles-mêmes ou dans leurs symbiotes microbiens de produits chimiques qui peuvent être utilisés pour contrôler les virus , les bactéries , les tumeurs et les champignons.
Dépourvues de toute enveloppe protectrice ou de moyens de fuite, les éponges ont évolué pour synthétiser une variété de composés inhabituels. L'une de ces classes est celle des dérivés d'acides gras oxydés appelés oxylipines . On a découvert que les membres de cette famille avaient des propriétés anticancéreuses, antibactériennes et antifongiques. Un exemple isolé des éponges Plakortis d'Okinawa , la plakoridine A , a montré un potentiel en tant que cytotoxine pour les cellules de lymphome murin.





.jpg/1280px-Euplectella_aspergillum_(cropped).jpg)